Bacteria (plural of late Lat. bacterium, Gr. baktērion, dimin. of baktron, 'stick,' 'staff'). It is a familiar fact that if water which contains or has contained organic matter either of animal or vegetable origin be left to itself, it soon becomes cloudy and covered with a film. If the floating flakes or surface scum be examined with the high power of a microscope, a marvellous exhibition of the lowest forms of life is presented. Representatives of the simplest animals, such as Infusorians, will probably be seen, and in some cases also some of the lowest single-celled plants, but all these will appear as giants amid a crowd of dwarf organisms, most of which are Bacteria. These usually appear as minute spheres, rods, or threads, propelled along by delicate lashes, or quivering together with the usual tremulousness of very small floating particles, or lying more passively entangled in a jelly-like flake. In a short time it will be seen that these bacteria multiply with extraordinary rapidity by cross division; one soon becomes a thousand, and the minute specks which first appear may, if the water contains enough food, soon form a mass filling the vessel. Wherever organic matter is in process of decomposition, in infusions as above indicated, or in disease or death, or within the living and healthy organism, there these bacteria are to be found.
So abundant are they, that in spite of their minuteness they did not escape the enthusiastic observation of the early workers with the microscope. Thus, in the 17th century, and with the comparatively imperfect microscope of those days, Leeuwenhoek described some of these low organisms. In 1773 O. F. Müller established two genera, Monas and Vibrio. Not much progress was made, however, till about 1838, when Ehrenberg and Dujardin included bacteria in their investigation of minute organisms. They referred the forms which they described to the animal series among Infusorians, and united a large number under the general title Vibronia. Nor was this reference surprising. At that time naturalists had hardly begun to realise that animals had no monopoly of mobility, though it is now a commonplace observation that the simplest plants exhibit a power of locomotion which is almost wholly lost in the higher forms. The argument in favour of the animal nature of bacteria which was once furnished by their movements is thus no longer specially cogent. That was not all, however. It is well known that the vast majority of plants have a characteristic way of feeding—deriving their carbon from the carbonic acid gas mixed with the air or water, and their other food-products from inorganic substances in aqueous solution. They take up the matter necessary for nutrition in a much simpler form than is possible for animals. This they are able to do as regards the carbon through their possessing the power of making a green colouring matter known as chlorophyll, by the aid of which the carbonic anhydride is absorbed and split up in sunlight. Animals, on the other hand, are only able to utilise food-products which have already been lifted up to a comparatively high level, which have in fact been worked up by plants or by other animals into complex compounds known as albuminoids—fats, starch, sugar, and the like. They cannot directly feed on the carbonic anhydride of the surrounding medium, nor on inorganic substances in solution in water. An appreciation of this physiological difference between plants and animals led to some difficulty in regard to the position of forms like bacteria. For in the first place they have no chlorophyll wherewith to utilise carbon at the low level of carbonic acid gas, and in the second place the nature of their occurrence plainly shows that they feed on the products of decomposition of plants or animals—physiologically speaking, very much as animals do. But as the forms and life-history of the simplest organisms were gradually elucidated, it became evident that such physiological distinctions were not only not hard and fast, but were delusive as a basis of classification. It was seen that though bacteria and other lowly organisms now known as simple fungi did not feed as the majority of plants do, yet they were in structure and development so entirely like unmistakable vegetables—the simple Algae—that separation was impossible. To Colin (1853) is due the merit of having established, on grounds of structure and life-history (morphology and development), the fact that bacteria are plants, and all succeeding research has confirmed his conclusion, which Nägeli (1857) corrected in detail, however, by referring the bacteria or Schizomycetes not to Algae, but to the parallel class of Fungi.
The bacteria or schizomycetes may thus be defined as extremely small, single-celled fungoid plants without chlorophyll, remaining single or united in loose colonies, reproducing rapidly by cross division or by the formation of spores, often occurring in myriad crowds enveloped in a jelly-like secretion, or separately with the power of energetic movement, and almost always associated with the decomposition of albuminoid substances.
Terminology.—The term micro-organism is convenient as a general word, especially when the position of the form in question is still undetermined. The French word microbe, introduced by Sédillot in 1878, has been approved by Littre, and is very widely used. It means a small living organism, and was invented to avoid dispute as to the plant or animal nature of low organisms like bacteria. It is now virtually equivalent to the latter. Bacterium is, like bacillus, properly the name of a distinct genus of schizomycetes, but the generic titles have both been too indiscriminately bestowed to be of much use, and are better restricted to popular designation. The term bacteria will be used throughout this article as equivalent to the technical title schizomycetes.
The physiological significance of bacteria in relation to disease and fermentation will be the special subject of the article GERM THEORY. The present article has to do with the following points only—(1) distribution; (2) form and structure; (3) life-history; (4) general classification; (5) more important forms; and (6) the methods of research. For further details as to position and classification, see SCHIZOMYCETES.
I. Distribution.—Bacteria are found practically everywhere—in air, water, and soil; in the mouths of men as well as on the walls of their houses; on the hair of the head and the toes of the feet; in chalk and coal; in food and drink; but especially where there is disease, death, or decomposition. To speak of the 'omnipresent bacillus' is hardly an exaggeration. (a) Air.—In the observatory of Montsouris, in Paris, M. Michel has charge of a special laboratory for investigating the times and seasons of atmospheric germs. In Britain also, some attention has recently been directed to the prevalence of micro-organisms in the air of various regions—lodging-houses, school-rooms, sewers, &c. In a cubic metre of Montsouris air there are on an average eighty bacteria. The maximum abundance is in autumn, the minimum in winter. The number rises in drought, and is reduced by heavy rainfall. Winds from crowded quarters, hospitals, slaughter-houses, &c., float a large number of microbes to the observatory, but pure air from lofty regions was found to contain hardly any. Sunlight is probably the most effective antagonist of these subtle enemies.
Some bacteria can only live in presence of atmospheric oxygen, and these are termed by Pasteur aerobic; others derive their oxygen from the organisms or decomposing media in which they live, and these are known as anaerobic. (b) Water.—Even pure water contains many bacteria, and always more than occur in air. The minimum occurs in condensed water-vapour (900 to the litre), the maximum (80 millions per litre, pint) of course in sewer water, especially if there be the least stagnation. If there be a proper flow of water, however, the germs never become dry enough to be floated in the air, and are not therefore dangerous. The sulphurous springs of the Pyrenees contain an abundant bacterium (Beggiatoa), which accumulates sulphur in its cell, and is especially abundant in the floating scum known as glairine or barégine (see BARÉGINE). Some bacteria, often called chromogenic (or colour-making) on account of their bright pigment, occur in water under certain conditions, and have given rise to superstitious accounts of 'blood-rain.' The red colour of stagnant pools in autumn has been known for fifty years as the result of a micro-organism, described by Ehrenberg as Ophidomonas sanguinea, but now known as a species of Spirillum. It rapidly changes from green to red, and if a waterspout draw up and re-discharge the bacterium-laden water, a shower of 'blood-rain' is no longer mysterious. Snow is sometimes coloured in a varied way by the presence of a similar organism (Micrococcus); which is not, however, to be confounded with the much larger Protococcus of 'red snow.' (c) Soil.—Nor is soil free from bacteria. Spring-water fresh from the source carries a cargo of these micro-organisms. Pasteur found that the germ of splenic fever (Bacillus anthracis) occurred in great abundance in the earth round about the pits in which diseased cattle had been buried. He also found bacteria in the earthy excrement of worms. Some refer the origin of most epidemic diseases to the ascent of these deadly germs from the soil, and if this be in any way true, the danger of drying marshes, of narrowed river beds, and of dust generally, is obvious enough. Some striking researches by M. Béchamp go to show that bacteria may lie dormant in soil for an incalculable period. A fresh piece of chalk taken from the quarry, with due care to exclude extrinsic germs, gave rise to abundant bacteria. He also discovered their presence in coal, but was unable to bring the sleeping germs into life again. Micro-organisms of modern date have also been found abundantly by M. Parize in the bricks of walls, and others have been found in association with the formation of salt-petre in the soil. (d) Food, &c.—The rancidity of butter, the putrefaction of cheese, the game-flavour and high odour of meat, the yellowness and blueness of milk kept in imperfectly scalded vessels, the excessive staleness of bread, and a crowd of other unhealthy conditions in food, are largely due to the presence of bacteria. Bloody stains on bread, meat, paste, &c., have also been traced to the presence of a brightly coloured micrococcus. In the same way the bitterness, ropiness, &c. of bad wine are due to the same organisms. (e) Decomposition and Disease.—A large number of bacteria are known to occur in direct association with pathological processes of decomposition in plants and animals, without apparently having any direct connection with the decomposition. These are Saprophytes (q.v.), like fungi generally. Others, however, have been proved in many cases to be the causes of pathological conditions in men and animals, a profound fact fundamental to the Germ Theory (q.v.) of disease. In contrast to the sapro- phytic bacteria, these are conveniently described as pathogenic. It must not be supposed, however, that the presence of a bacterium in an organism is necessarily associated with abnormal decomposition or with disease, since some occur quite normally, and without any evil influence.
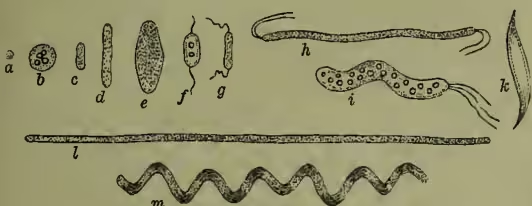
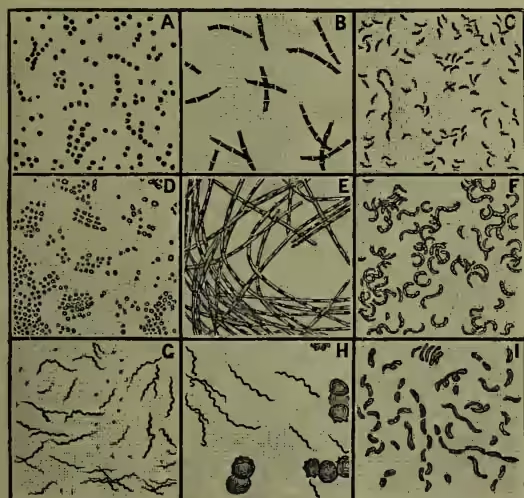
II. Form and Structure.—As regards the shape of the individual units, four principal forms may be distinguished—viz. spherical, elliptical, rod-like, and spirally curved. It must be noted, however, that a bacterium may, as Lister and others have shown, pass from one form to another in response to different physiological conditions, that a species spherical in its young stages may be elliptical or cylindrical afterwards, or that a rod-like form, such as Bacterium lactis, which causes lactic acid fermentation, may become thread-like or spiral when sown in urine.
Bacteria do not, however, remain single, but reproduce by division, and the results of division may remain loosely united. (1) The spherical micrococcus and the elliptical or cylindrical bacterium always divide in one direction, and the resulting couple may remain temporarily united, or continue multiplying to form a myriad colony embedded in a jelly (zooglea). (2) In Bacillus, Leptothrix, and Beggiatoa, the cells may form long cylindrical threads, and this is varied in Vibrio, Spirillum, and Spirochaete by the development of spiral or wavy curvature. (3) In Sarcina, again, the division takes place in three planes, and the result is obviously the formation, not of long chains, but of cubical clumps. Just as among simple algæ, so here the results of division form—(1) isolated spheres; (2) longitudinal or even branching filaments; and (3) planar expansions, always, however, extremely minute.
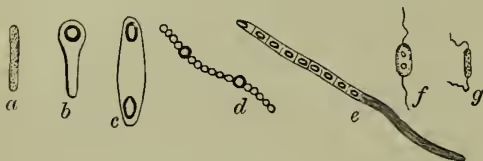
Structure.—The unit mass of protoplasm which forms the bacterium individual is always inclosed in a membrane, which sometimes consists of the cellulose characteristic of the cell-walls of plants, but more frequently of a peculiar albuminoid substance which has been designated mycoprotein. This membrane may be stiff or flexible, colourless or brightly pigmented, and undergoes uniform increase in thickness. The contained protoplasm consists largely of the above-mentioned mycoprotein, and may include fat-like granules, particles of pure sulphur, grains of starchy material, and dissolved pigment often of a brilliant colour. No nucleus has yet been detected. In all bacteria, except those which form long filaments, locomotor organs are present at some stage or other, as delicate lashes or cilia always situated terminally. These have been oftenest observed in the young swarm spores, and 1–4 may be present. In the adult forms they apparently arise when the protoplasm is needing oxygen, like the long tongues in a dying flame, and they serve to hurry the bacterium to the surface or to some position where oxygen is more abundant.
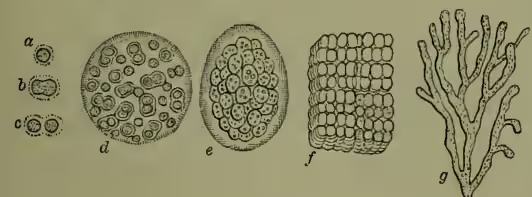
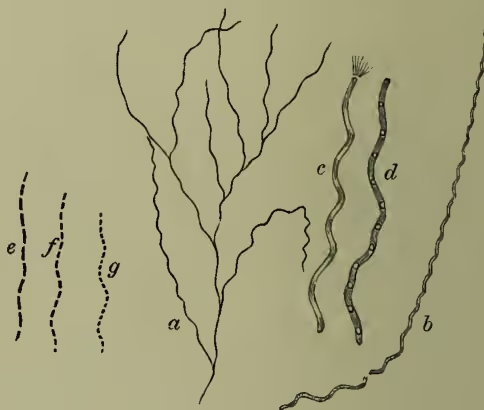
III. Life-history.—It is convenient to distinguish in the life-history three important events or processes—(a) increase in size and modification of form; (b) reproduction; (c) the assumption of the resting form known as zooglea. (a) Increase in Size and Modification of Form.—As the result of abundant nutrition, the bacterium unit increases in size, and this growth necessitates division, and is associated with change of form. The minute spherical cocci usually grow into rod-like or cylindrical shapes, and these little rods divide rapidly, ranging themselves in rows to form filaments. In those filaments the original distinctness of the component units is often lost. Nutritive and other conditions, however, affect the form, both of the separate rods and of the composite filaments. Both exhibit a tendency to be more or less spirally curved. Thus wavy forms arise from the longer forms arising from rows of straight rods are really multicellular, whether they appear to be so or not. This modification of form is technically known as the pleomorphism of bacteria. The various forms—filamentous, incipiently branched, and plane—which are exhibited as the results of multiplication, have been already referred to. One other fact must be noticed, that the composite chain of bacterium units never remains perfectly uniform throughout. The parts of the chain are not all in exactly the same physiological conditions, and variations therefore occur abundantly. The end of the thread may become much enlarged, or here and there individual masses may preponderate in size over their neighbours. These variations offer interesting parallels to those occurring in filamentous algæ. (b) Reproduction.—The more complete the degree of parasitism in fungi, the more probable is the absence of sexuality. In bacteria, the multiplication is an entirely asexual process. The absence of anything of the nature of sexual union may perhaps in part explain the infinitesimal minuteness of the individuals; and again, since the bacteria live to a certain extent bathed in waste products, the stimulating character of the medium has been regarded as a physiological substitute for the stimulus to division usually supplied by fertilisation. The process of multiplication which invariably occurs is by means of division. The coccus or rod-like form becomes somewhat elongated, a double partition wall is developed in the middle, and the two daughter cells are thus formed. If the results remain united, and if the lines of division be all in the same direction, the filaments above mentioned must arise. The resulting forms recall species of Nostoc and other Algæ. When the division occurs in more than one plane, the result is the formation of plates as in Bacterium merismopedioides, or of packets as in Sarcina ventriculi, or of discs as in Crenothrix.
Besides this external and successive fission, another process is frequently observed in which the divisions occur in a less definite way. Spiral or other forms are seen to divide rapidly into fragments, either at once externally, or at first only internally. The rupture of the flexible filament occurs in the more passive middle portion, and each half again splits, and so on. The liberated fragments grow again to the adult size.
But in addition to these entirely vegetative processes, bacteria also reproduce by means of special cells—the spores. The contents of the units contract, become rounded off, and surrounded by a membrane. It is a special case of internal (endogenous) division, and has been observed in cocci, rods, and spiral forms. In some cases, such as Clostridium butyricum, of butyric acid fermentation, the cells which will give rise to spores can be distinguished from the others before the spores have begun to appear. Spore-formation appears to set in as a response to insufficient nutrition, as is the case in very many instances of reproductive activity. The liberated spores germinate and develop into the adult forms. Often ciliated in their young stages, they become more quiescent with increasing size. (c) Formation of Zoogloea.—Wherever bacteria have been allowed to flourish unmolested, jelly-like flakes or clumps are found. These were formerly regarded as distinct forms and called zoogloea, but they are now recognised as a phase in the life-history. Such gelatinous clumps are often conspicuous both in size and colour; thus that of Clostridium polymyxa may measure more than an inch across, while the 'frog-spawn' zoogloea occurring not unfrequently in the beetroot juice used in sugar manufacture, may attain a size of more than a foot. These zoogloea consist of myriad colonies of bacteria embedded in a jelly. Their formation is due on the one hand to the aggregation of resting bacteria, arising from one mother cell or from several, and on the other hand to the thickening and gelatinisation of the individual membranes. The same habit of aggregation is not uncommonly exhibited by simple unicellular organisms, and also by the cells of higher animals, though in these cases the individual units fuse more or less completely to form a composite mass or plasmodium. Nor is the gelatinous degeneration of the membrane in any way unique, but is exhibited both by algae and fungi.
IV. Classification.—A thorough classification of bacteria has yet to be elaborated. The multitude of forms differing in comparatively trivial points of structure, the insufficient state of our knowledge of the life-history of many genera, the difficulties involved by the abundant pleomorphism, the existence of weighty physiological differences between forms which seem otherwise absolutely alike, make a dogmatic classification at present quite impossible. In 1838 Ehrenberg distinguished four genera—(1) Bacterium—straight and rigid; (2) Vibrio—snake-like and flexible; (3) Spirillum—spiral and rigid; (4) Spirochete—spiral and flexible; while Dujardin united the two last into one genus. In 1872 Cohn, to whom so much progress in bacteriology has been due, distinguished four dis- tinct tribes—(1) Sphærobacteria—globules (Micrococcus); (2) Microbacteria—short rods (Bacterium); (3) Desmobacteria—long rods (Bacillus and Vibrio); Spirobacteria—spirals (Spirochete and Spirillum). This classification held ground for a few years, but has been abandoned for a reason which must already be obvious. In 1873 Lister was the first to hint at the fact of pleomorphism. He showed that certain forms referred to different genera and groups were really phases in one and the same life-cycle. The mistake had been made of separating young and adult stages, and of regarding as permanent and fundamental shapes and habits which were only temporary and transitional. Till the life-history of all the forms is completely known, the same mistake in this, as in other departments, is sure in some degree to persist. What Lister suggested was in the same year even more conclusively demonstrated by Lankester. Forms belonging to different groups of Cohn's classification were shown to be successive chapters in the life-history of one species. The researches of Billroth (1874), Klebs (1875), Nägeli (1877), Warming, and others, but most of all, perhaps, of Zopf, have established the prevalence of pleomorphism, and have made this fact at least certain, that whatever the final classification is to be, it must be one which takes account, not of specific facts of form, but of the whole round of the life-history, and of the sum-total of morphological and physiological properties. Any classification especially of lowly organisms, where the range of differentiation is always comparatively slight, ought to take account particularly of the two planes of development—vegetative and reproductive (see ALGÆ). But as we have seen, the range of vegetative modification is very limited in bacteria, and the same is true of what of the reproductive processes is now known. De Bary has, however, suggested the distinction of two great groups—those which form spores (endospores), and those which become divided into segments which are equivalent to spores, but are not internally formed (arthrospores). With further knowledge of the reproductive processes, the classification will be gradually elaborated. Two other points must not be overlooked. Recent studies on fungi and algae, which differ so markedly in the respective absence and presence of chlorophyll, tend increasingly to emphasise the close correspondence in structure and life-history between different groups in these two classes of plants. Instead of there being two separate branches in the genealogical tree, one representing fungoid, and the other algoid forms, it has been repeatedly suggested that the various subdivisions of the fungi are to be regarded as the chlorophyll-less representatives of parallel subdivisions among algae. This must be kept in view in regard to bacteria. Before proceeding to note the most familiar classification and some of the more interesting forms, it must be again observed that a few of the generic titles have come to be used in very vague and ambiguous ways. This is so conspicuous in regard to the terms Bacterium and Vibrio, which have been used so widely that some authorities incline almost altogether to suppress them as accurate generic designations. The provisional classification now most commonly adopted is that of Zopf, as expressed in the following table. He distinguishes four groups—(1) Coccaceæ; (2) Bacteriaceæ; (3) Leptotrichæ; (4) Cladotrichæ.
(1) Coccaceæ.—Only cocci, singly or in contact. Division in one or more directions.
(2) Bacteriaceæ.—Mostly with cocci, and also rods (straight or bent), and thread-forms (straight or spiral), without distinction between base and apex. Division in one direction.
(3) Leptotrichæ.—Cocci, rods, and thread-forms (straight or spiral), with distinction between base and apex.
(4) Cladotrichæ.—Cocci, rods, threads, and spirals. Thread-forms with false branches.
V. Important Forms.—(1) Coccaceæ.—Streptococcus is a genus with numerous species, some associated with disease in men and animals—e.g. with diphtheria, yellow-fever, foot-and-mouth disease, others merely feeding on the results of pathological processes, and a few entirely unassociated with diseases of animal life. Cubes or packets of Sarcina are found forming coloured patches in various situations. Micrococcus has been observed in cases of scarlatina, measles, whooping-cough, typhus, &c., but their precise rôle is not certainly determined. Hydrophobia is believed by many to be due to the presence of a micro-organism, and micrococcus has been observed in this connection. simply saprophytic, following and not causing pathological processes. M. amylinovor has been described as the cause of 'fire-blight' on pear-trees and other plants. Many of these forms are brightly coloured, as, for instance, that species which causes the blood-red sweat in the human armpit and elsewhere.
(2) Bacteriaceæ.—Species of bacterium have been found associated with pneumonic disease, diphtheria, &c.; others cause fowl-cholera, pebrine of silkworms, and other diseases of animals; and a large number have been described apart from any directly pathological connection. Bacterium prodigiosum forms blood-red zoogleea, and occurs on bread, paste, milk, and such commodities. Its appearance has given rise to superstitious anecdotes about 'blood-rain,' and the like. In 1843 this fungus was so prevalent in Paris that it caused a sort of bread-plague, especially in the military bakehouses. B. aceti oxidises the alcohol of wine and other fruit-juices into vinegar. B. termo, which is so often described as an almost omnipresent organism, is apparently only a phase in the life-history of many different forms. A species of Spirillum is believed to be the cause of relapsing fever, and Koch has described another form—the Comma bacillus—which he has found associated with Asiatic cholera.
A large number of species are known apart from disease. The frog-spawn fungus, otherwise known as Leuconostoc mesenteroides, is of some importance, since it sometimes invades and corrupts the beet-root juice and molasses used in sugar manufacture. Bacillus is a large genus with numerous formidable species associated with the diseases of men and animals. Such are the bacillus of leprosy, syphilis, and typhoid fever, tuberculosis, splenic fever, glanders, swine fever, &c. The bacillus of 'blue milk,' of hay-infusion, and Bacillus septicus of putrid albuminous fluids, may be noted as examples of forms unassociated with disease. A species of Clostridium is the cause of the disease of cattle known as 'black leg' or 'quarter-evil.'
(3) Leptotrichæ.—A species of Crenothrix with long filaments is sometimes abundant enough to stop up narrow water-pipes. Beggiatoa occurs in various forms in sulphur springs, on sea-water, on the surface of marshes, &c. The best-known species has a peach-blossom red colour. Leptothrix buccalis occurs abundantly in the mouth, and is apparently associated with the decay of teeth.
(4) Cladotrichæ.—As a representative of this last group of bacteria, Cladothrix dichotoma must be noted, a form with false branches, occurring more abundantly than any other in water containing organic matter.
VI. Methods of Research.—Besides the usual apparatus of any well-equipped laboratory for the study of minute structures and organisms, a number of special appliances are required for the successful investigation of bacteria. Thus, since the intrusion of germs other than those which are the specific object of research is a constant danger, there must be some means for sterilising the tubes, tools, media, &c. This is generally done by means of a steam or hot-air steriliser, in which all the extrinsic germs are killed off. Incubators are also used for purposes of cultivation. The bacteria obtained in endless ways may be examined as they are, or stained with reagents to bring out the individual structure, or since the life-history is all-important, left to grow, and watched at their successive stages. They used to be left in some sterilised fluid, such as broth, blood-serum, urine, milk, or Pasteur's fluid, and allowed to grow in test-tubes, or other vessels, plugged with cotton-wool. It is, however, exceedingly difficult to get a perfectly pure fluid medium, nor was it possible in such cases to isolate
The researches of M. Pasteur have thrown light on this problem. Many micrococci are apparently the different kinds of bacterium which might be present. In view of this, Koch has recently introduced the method of cultivation on sterile solid media. Sterile nutrient gelatine, or some such substance, is liquefied in a tube and inoculated with the bacteria in question. These are distributed through the fluid, which is then poured out on a plate of glass and left to solidify. The various bacteria can no longer move about and mingle with one another, but are fixed to one spot, where they develop. The resulting fixed colonies can thus be studied without confusion. Slices of sterilised potatoes are also very frequently used as solid media for the cultivation of bacteria. Finally to elucidate the relation of a micro-organism to a given disease, it is necessary not only to have obtained it from an organism suffering from the said disease, but it is imperative that some of a pure cultivation be introduced into a healthy organism, to see whether it does or does not cause the disease. The inoculation may be brought about by inhalation, or along with the food, or by injection in some form or other.
How such methods have, in the hands of investigators like Pasteur and Koch, resulted in discoveries of profound importance, not only to a scientific theory, but to the practical amelioration of life, will be discussed in other articles.
See ALGÆ, ANTISEPTICS, FERMENTATION, FUNGI, GERM THEORY, INFECTION, SCHIZOMYCETES, &c.; also Zopf's monograph on Bacteria, Die Spaltpilze (Breslau, 1885); De Bary's Lectures on Bacteria, (2d ed. Leip. 1887); Klein's Micro-organisms and Disease (1886); Löffler's History of Bacteriology; and manuals of Bacteriology by Trouessart ('Int. Sc. Ser.' 1886), Crookshank (1887), Sternberg (New York, 1892), Sims Woodhead (1892), and Günther (3d ed. Leip. 1893).