Vegetable Physiology. The seasonal sequence of leaf, flower, fruit, and seed ever reminds us that plants live and change; and the cycle of farm or garden operations is correlated with this. The relation of these processes to plant development must be closely and carefully examined, and the manifestations of life seen as the resultant of many hidden internal actions. It is useful to begin by sowing some seeds and studying the conditions under which they grow best, and what changes constitute this growth, and on these germination experiences base a fuller investigation of the life-processes of the adult plant.
Germination (see SEED, SPORE, &c.).—The seed sown must be alive. The common test—that a living thing breathes—may be adopted. In many cases this Respiration (q.v., and see also below) can be shown by drawing the air surrounding the seeds through a tube containing lime-water, when the insoluble white carbonate of lime is formed by the union of carbon dioxide, CO2, with lime. In some cases transpiration (see below) can be detected. Most seeds in temperate regions live a year at least, sometimes two or more. Where there is no winter, some seeds (as those of the coffee) must be planted almost at once. On the other hand, melon seeds have germinated when over forty years old. The young plant must be nourished by food stored around or within it, until such time as it can make its own food. These reserves (see ALBUMEN, SEED) are not directly assimilable by the embryo, but must first be digested by 'ferments' (see FERMENTATION, and below), which exist in necessary quantity only at a certain period of the seed's life. The reserves are often very dry, and must have a supply of moisture before digestion and assimilation are possible.
No motion of particles bringing this about can take place if the seed be frozen; so a minimum of heat—from 4° to 13° C.—is necessary, and germination is impossible at temperatures higher than 36° to 46° C., and is most active at 26° to 35°, varying according to the nature of the seed. Other external influences are of comparatively little importance. The visible results of germination are the appearance of root, seed-leaves, and shoot. The life-processes of the seed have all been intensified—breathing is more rapid, transpiration is marked, and sometimes enough heat is given out to let us feel and measure it—e.g. in malting. The seedling is very sensitive to all outside influences: it quickly droops in drought, shrivels up should the sun's heat be too great, and blackens if the nights be frosty. As the plant develops it becomes less influenced by environment, and soon begins to make its own food; but before discussing this special function of plants it is well to look at the life-processes common to all organisms.
Respiration (q.v.) is manifested by the intake of oxygen, O, and the final liberation of carbon dioxide, CO2, gas. Attempts to grow most plants without free O are failures; and the CO2 can be precipitated as calcium or barium carbonate when drawn through lime or baryta water. The activity of respiration is best measured by the number of volumes of gas per unit mass taken in and given out in unit time under constant conditions. Thus comparative accounts of the rate of breathing of different plants and of different parts of plants may be obtained, as well as the relative intensity of respiration at different ages of organism and of organ. Succulent plants and very dry ones do not respire so energetically as rapidly growing grasses and annuals; and the young plant breathes very actively, whereas the hibernating bulb or seed respires most sluggishly. External influences have no great effect on respiration, which, however, is somewhat quickened by a rise of temperature up to that which kills the plant. For most plants in normal conditions the volume of oxygen taken in is approximately the same as that of CO2 given out.
Most cells are not immediately asphyxiated if deprived of free oxygen, but take it from some of the simpler organic compounds they contain or that surround them—a leading product in many cases being Alcohol (q.v.; and see FERMENTATION). The yeast plant has been domesticated for this purpose, and other moulds and also many bacteria are able to prolong the process and convert large quantities of sugar into alcohol, or split up other substances into different constituents. Here the volume of CO2 exhaled is greatly in excess of the O inhaled. The volume of O consumed is much greater than that of CO2 liberated in other cases, such as that of the acetoic ferment.
Transpiration.—The heat given off in respiration is partly used in evaporating water, ever present in organic substances. This vaporising of water is usually termed transpiration. It may be shown by placing a plant in a pot on one of the scales of a balance, and noticing that the weight lessens, although, as we shall see, a green plant in the light must be adding to its substance all this time. Accurate estimations of the volume of transpired water can be made by sealing the moist soil and the root of a plant in a glazed pot covered with a waterproof lid, placing this under a bell-jar, supplying the plant with water-free air, and measuring the change of weight of the drying tubes through which this air is sucked after passing through the bell-jar.
The amount of water vaporised is by no means constant. It varies with nature and age of plant, and part of plant, and with external conditions. Some seeds, especially oily ones, cannot be found to transpire, others can; young plants readily do so, and leaves more vigorously than roots or stems. Transpiration seems very analogous to evaporation, and is less marked in moist conditions, but it never ceases in saturated air as the latter does. It is said to be less than evaporation, because a plant killed and uninjured (by anaesthetics for instance) gives off more water vapour after death than before it; but the experiments are inconclusive. In fungi and other greenless plants the influence of light is noticeable to a slight extent, increasing transpiration. But if a green plant be kept in the light there is a much stronger marked maximum when the sun is brightest. When this plant is kept under a sealed bell-glass for some time no CO2 but an excess of O is found. If this last change be prevented by ether, respiration being unaffected, then the amount of water vaporised is increased. All these phenomena are closely related, and will be discussed below. The leaf is the green organ par excellence, and by its construction, its distribution on the plant, and its position with the surface normal to the sun's rays (except when these are excessive) it can arrest the greatest possible number of radiations (see LEAF).
The maturing leaf has two transpiration maxima. The first and greatest is when the leaf is very young and every surface-cell can transpire freely. Gradually a cuticle develops which may be coated with wax in addition, and then the transpired water passes by the stomata. The crescentic bounding cells of these openings are specialised, and contain Chlorophyll (q.v.), which, absorbing radiations, sets up activities synchronously with those of other green tissue of the leaf, and modifies the contents of these guard cells, so that they swell and open the stomata to their widest when the other green cells are most active. The second transpiration maximum occurs when most stomata are developed.
'Assimilation' and 'Chlorovaporisation.'—When a plant grown in the dark is brought into the light it first turns green, and then begins its new functions. The yellow etiolin stops certain ether waves which transform it into green Chlorophyll (q.v.). The spectrum of chlorophyll shows absorption bands in the red and violet, with less marked ones in the yellow. These waves move the molecules of plastic matter (protoplasm, solids, and solutions) associated with this chlorophyll, so that CO2 is taken up and O is ultimately set free. This 'assimilation' is the reverse of respiration, a deoxidising process, an unburning where the sun's rays supply the needed energy. It often masks the breathing, and early physiologists were led to conclude that green plants respire in the reverse sense to animals. The volume of O given off is approximately the same as that of CO2 taken in. Naturally optimum conditions of light promote optimum 'assimilation,' and the minimum occurs at night; but too bright light is injurious. Heat, up to 30° C. at least, increases it, and any optimum must be above this. A young leaf 'assimilates' more vigorously than an old one. The increase of transpiration when a green plant is in the light is probably due to all the energy arrested by the chlorophyll not being spent in 'assimilation,' but in part vaporising water; and this seems probable not only because more water is transpired when assimilation is inhibited by anaesthetics, while respiration is unaffected, but it is found that the blue and violet rays absorbed are more efficient in transpiration, while the red ones are most active in assimilation. M. van Tieghem has termed the transpiration, which is a function of chlorophyllic activity, chlorovaporisation, and others transpiration chlorophylliène. Chlorovaporisation is a useful term when it is necessary to distinguish between it and the transpiration due to protoplasmic activity.
Absorption.—Plants living in water have all their epidermal cells absorptive, and need no elaborate water conducting system. Land-plants must either extract the water from the air, or from the soil, or from both. Experiment shows that the Roots (q.v.) are the active absorbers, and the fine root-hairs near the tip of the roots the special organs concerned in this. These absorbing hairs, as they are best termed, are drawn-out epidermal cells with walls of Cellulose (q.v.), differing from that of most cells in being more readily permeated by water, inside which is a lining layer of Protoplasm (q.v.). This double layered cylindrical bag is filled with cell-sap.
The contact between these hairs and the grains of soil is very close, and a film of water coating their outside is continuous with that adhering to the particles of soil. The sap is more viscid than this mineral water outside, and so we have a living cell acting as a complex dialyser (see OSMOSE)—colloid cell-sap, crystalloid soil water, with protoplasm membrane supported by permeable cellulose skeleton work between. We have thus the conditions necessary for a flow of mineral water into the plant (any outflow being very small, although when acid aiding in the decomposition of the soil), which is continuous as long as these conditions remain constant. The cell-sap made less colloid by this influx of water is crystalloid compared to that of the inner cell, and the current accordingly flows inwards. Yet this is but the crudest approximation to a full explanation, for not only are the variations of the cell-sap's quality due to this dilution to be considered, but also those due to protoplasmic activity, and in addition the fact that the osmotic properties of the protoplasmic membrane itself are ever being modified by its continual oxidation and deoxidation.
Movements of Water.—In water-plants and those living in damp conditions, as well as in low-growing plants, osmose is the main agent. We need not imagine a long series of cells, each with its sap more colloidal than that of the next in order; for even in the laboratory the reversing of a membrane sometimes alters the direction of the current, and the plant membrane is very complex in structure, as well as constantly changing. It is difficult to set a limit to the influence of osmotic action.
Experiments making plants absorb tinted water show that it passes up the wood (xylem) part of bundles (see STEM), as may also be proved by noticing that only when the young wood has been incised all round do the leaves wither, and even hollowing out old wood does not induce drooping. Conifers have true vessels only in first-formed wood; hence any satisfactory explanation must not lay too much stress on capillarity, which would account for a rise of a few feet at most. Much stress has been laid on the porosity of the cell-walls and their permeability by water, and Sachs in his 'imbibition theory' has emphasised the mobility of this water, and believes he can explain the phenomena by its aid. The transpiration of water helps to raise water owing to pressure differences resulting from it, and these may be increased by the bubbles of air in the vessels or fibres losing their oxygen to respiring cells, the given off being dissolved in the water. But pressure differences can account for no more than a few yards of rise.
The pressure from below at some periods of the year is very considerable, as Stephen Hales showed in his classical experiments on 'root-pressure,' when the watery sap was forced up a tube tightly fastened to the end of a cut stem. This is most noticeable in spring, and in part at least is a function of temperature, for a slight heating of the soil will sometimes make an experiment work more rapidly. This 'root-pressure' gives rise to 'bleeding' when the sap is forced out at buds in spring, and to 'dew' when exuded along the edge of a Lady's Mantle or other leaf. This exudation occurs from stomata in grasses, &c., and from special organs in saxifrages, balsams, &c. (called water-pores or stomata). Should the water have passed through a sugar-laden tissue it becomes nectar, and the organ a nectary. Some plants show an excessive development of water-pores and nectaries, and may be specially modified so that the exuded sap may be retained—e.g. the pitcher-plant (see INSECTIVOROUS PLANTS).
Metabolism (q.v., and see PROTOPLASM).—We have seen how green leaves are supplied with gases, water, and dissolved salts, and how they can trap special ether waves. The active energy of these waves is used to transmute the simple inorganic compounds into complex organic ones, which in the process of respiration are reduced to simpler substances again, and the potential energy transformed into kinetic. These metabolic changes take place in living cells full of intense activities. Currents course through the protoplasm and cell-sap in every direction and between the cells, which are also united by strands of protoplasm. The gases used and given off in respiration and 'assimilation' are floated in and out, and each protoplasm particle burned or unburned is the centre of an area of disturbance. Pure protoplasm is influenced equally by all rays: that associated with chlorophyll is affected by certain red and violet rays in particular. These, especially the red ones, bring about the dissociation of the elements of the , the assimilation of the C, and the excretion of the O. But carbon is not found isolated in the plant. The simplest carbon compound found in a green leaf after a few minutes' exposure to light is starch. Remembering that the same volumes of are absorbed as of O excreted, we may write a chemical equation expressing results though not processes: . But we suspect that starch has had a more complex life-history, and that the first-formed organic compound is probably simpler, though very difficult to detect. Probably formic aldehyde is the product, and again the first and last chemical conditions may be written . From this higher compounds of C, H, and O may be formed.
Since the synthetic process is hidden from us, it will be best to give an idea of the gradual increase of complexity of the compounds found in plants. With nitrogen added to carbohydrates, Amides (q.v.) are formed, and when S and P are combined with these, the Proteids (q.v.). Protoplasm may be either a peculiar unstable arrangement of proteid molecules or a collection of many compounds in a similar unstable condition, so that the wandering oxygen molecules coming in contact with them explosively seize hold on some of the C, and completely unite with it in the most stable way. The protoplasm is constantly oxidising to be deoxidised again, burning to unburn. Irritable and unstable, it is formed to be destroyed, and recomposes only to be once more decomposed.
The result of this burning of protoplasm is either to form proteids, which may become protoplasm again or decompose into simpler compounds; or else to form a combination of compounds—proteids, carbohydrates, &c., which wholly or in part may form new protoplasm, be reduced, or remain in their new form. This may be conveniently summarised in fig. 1, A. The contrast between protoplasmic and chlorophyllic processes is shown in fig. 1, B. It is very difficult to discover whether a compound is a synthetic or a decomposition product; and it is just as hard to obtain some substances at all, since they are transferred to another cell, or transformed to another shape almost as soon as they are formed.
Fig. 1.
Other substances, however, may be stable for a long time. Any matter used to form protoplasm is called a food or reserve, and what is not utilised in this way is a waste. Both may be temporary or last for a considerable time. The same substance may be both a temporary and a seasonal reserve; for instance, starch found in green leaves during the day disappears at night; and starch is the main reserve in many seeds, tubers, &c.
The matter elaborated in the leaves is conveyed to growing and storing organs. Many constituents of elaborated matter cannot be transferred as they are, but must be changed to soluble substances, capable of passing through the cell membranes. This is effected by digestives, often called 'soluble ferments' (see FERMENTATION), which are found in all living cells. Insoluble starch is acted on by diastase, and changed to a cane sugar, which in turn is 'inverted' to soluble glucose; insoluble proteins are changed to soluble peptones by pepsin, &c. This soluble matter is transported by bast cells (see STEM), as is shown by peeling the bark off a tree, when growth takes place above the ringed part alone. A chain of living cells may osmotically conduct this elaborated sap, while the sieve vessels of the bast are active according to some, or according to others may be storehouses supplying food to the actively growing and dividing embryonic tissues (cambium) rather than conductors. Some wastes are at once excreted—e.g. O, CO2, H2O; some are secreted, and others are deposited in aggregates or crystals. Some so-called wastes are used again, and really are seasonal reserves. Many wastes are of great use in industry and medicine (see PHARMACY, NARCOTICS, GUM, RESINS, &c.). The odours and colours of plants are probably wastes (chlorophyll itself is sometimes thus classified).
Composition of Plants.—The plant uses O in respiring and CO2 in assimilating, both of which come from the air. Water and the salts dissolved in it supply the other elements which are absorbed by the roots. All substances washed out of the soil are not used by the plant, but cease to enter when its lower cells become saturated; whereas the constant assimilation of any useful salt makes it be constantly absorbed. Some plants use more of one substance than of another, and a soil soon becomes poor if the same crops be constantly grown on it.
Thus we need to have Rotation of Crops (q.v.) and also fertilisers (see MANURE). Plants may be grown in clean sand, and watered with nourishing solutions. But soil is not essential (witness lyacinth culture in our rooms), provided the roots are properly protected, supported, and supplied with water and the necessary minerals. An excellent solution for watering or growing plants is the following 'normal culture solution': Dissolve 1 gramme nitrate of potash, gramme sulphate of magnesia, gramme sulphate of lime, and gramme common salt in 1 litre of distilled water, and add a trace of phosphate of lime and of any ferrous salt.

The results of growing a plant in this normal culture solution are very satisfactory if external conditions are kept favourable; and when these latter are the same for all plants an excellent comparison of the effects of the lack of each of the compounds may be obtained, as is shown in fig. 2. The plant's needs regarding matter may be accurately determined by analysing all the gases given 1, maize plant grown in 'normal culture solution' (see above); 2 had no sulphate of magnesia; 3 had no common salt; 4 had no sulphate of lime; 5 had no phosphate of lime; 6 had no nitrate of potash; 7 had no iron, and has become chlorotic; 8 was grown in distilled water; 9 was grown in normal solution, but in the dark. off during life, and on combustion at death, and all the ashes that are left. The most important elements are C, O, H, N, S, P, and other essential ones are Ca, K, Na, Mg, Cl, and Fe.
Growth.—After germination the root of the young seedling spirally winds its way down among the particles of the soil, turning away from them with its sensitive tip, seeking out the moister parts. Behind the tip it clasps anything it touches and thus anchors itself. As it grows older it contracts, making this anchorage secure. The shoot, often with arched head, shoulders its way upwards towards the light, twisting round more or less as it rises. Root and shoot grow in thickness as well as in length (see ROOT, STEM). The curves and movements of growing parts are discussed under PLANTS (MOVEMENTS OF). The rate of growth varies very greatly. Bamboos are said to lengthen 10 inches in twenty-four hours. Our cereal crops (36-60 inches high) ripen about twenty weeks after the sowing of the seed, and some trees are hundreds of years old. Growth is most active in spring, and elongation is greater at night than by day, and indeed varies from hour to hour—the
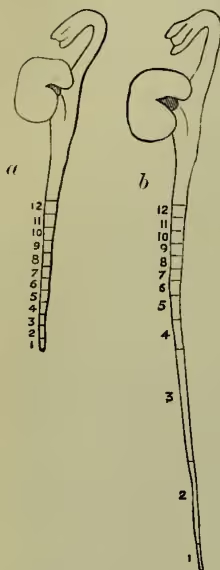
Victoria regia leaf, according to Drude's experiments, lengthening 99.4 mm. between 11 P.M. and midnight, and 127.9 mm. between 12 and 1 A.M. The greatest elongation takes place near the tip both of root and of shoot, within the last few millimetres in the former, on the last few centimetres in the latter. Fig. 3 represents the lengthening of equal divisions of a bean root after twenty-four hours' growth. and the energy from the heat given off in respiration. It is necessary to examine what changes constitute this growth more closely; but, unfortunately, a satisfactory mechanical explanation seems scarcely possible.
It is necessary to imagine all the stir and change going on in the Cell (q.v.). The protoplasm grows by new organic matter formed by the chlorophyll grains being digested and assimilated. At the limit of growth cell-division occurs (see CELL, REPRODUCTION). After a cell has ceased dividing, the mass of its protoplasm is probably not greatly increased, but vacuoles appear in it, and as the cell grows older and elongates the protoplasm finally forms a membrane bounding one large sap vacuole and lining the cell-wall. The protoplasm has been rapidly adding to the cell-wall, and at last the wall assumes its final shape and markings. The elongation is due to the distension (turgor) of the cell by accumulated water. When a plant is rapidly transpiring (for instance a green plant in sunlight) the cells are not so likely to become turgescent. This is one cause at least of the lengthening being greatest by night, and it is well to emphasise that it is elongation that is then a maximum, and not necessarily the substantial increase which makes it permanent. When the wall is stretched in this way new cellulose can most readily be added. The cellulose may be deposited as such, or precipitated from some compound the rest of whose constituents are soluble. The unit may be the cellulose molecule, or a more complex one formed of an intimate union of several molecules (micellæ of Nägeli).
There are several theories as to the way in which the cellulose in any of the conditions just mentioned may be deposited. It may be plastered on inside of the wall (apposition theory), or the protoplasm may penetrate the cellulose wall and there lay down new matter, or the cellulose may be floated into position between old particles (intussusception). The transformation of cellulose into wood (lignin), or cork (suber), or gum (mucilage) may be explained most readily by one of the two latter hypotheses. The constant passage of cell-sap, and the permeability of the cell-wall (unless corky or gummy) must never be forgotten.
Movement in Plants.—The constant currents in cell-sap and protoplasm may be co-ordinated so that the whole contents are caused to glide round and round the cell (Elodea), or stream from side to centre, and from the nucleus back to the wall again (stamen hairs, Tridescentia). Sometimes this affects the whole organism, which may creep along like an amoeba (Myxomycetes) or slowly swing to and fro (oscillatoria); or may be concentrated in one special part of the organism (cilium) whose rapid movements transport the plant from place to place (Volvox, zoospores). The sleep of plants may be explained as a partial periodic movement of plants, and classed among these natural movements. A number of motions are to be seen when a plant is specially stimulated, and these movements of irritability are discussed under PLANTS (MOVEMENTS OF).
Habits of Plants.—The simplest plants live in water, but other forms exist underground (Truffles, q.v.), while some can live in sugar (Yeast, q.v.) or in oil. The majority, however, are fixed to the soil and spread their expansions in the air. Some simply scramble along the ground (Brambles, q.v.), some keep erect, and others twine and climb up their stronger neighbours or other support to reach the light. Plants may live on decaying matter (Saprophytes, q.v.), or on other living plants (Parasites, q.v.): many associate with other plants or animals, each mutually helping the other (Symbiosis, q.v., or Consortism).
Reproduction.—At the limit of growth cell-division occurs; and sometimes special cells are cut off which either separately or after union with another cell are capable of reproducing the plant (see REPRODUCTION, FERTILISATION, &c.).
LITERATURE.—The works of Stephen Hales, Ingen Housz, SÉNÉBIER, De Saussure, De Candolle, Hofmeister, Boussingault may be consulted. The best books in English are Vines's Physiology of Plants (Cambridge, 1886), Sachs's Lectures on the Physiology of Plants, translated by Marshall Ward (Oxford), Goodale's Physiological Botany (Macmillan, 1890), and an excellent introductory book is Master's Plant Life ('Life on Farm' series). For experimental work Detmer's Pflanzenphysiologische-practicum (1888) is very useful. The physiological chapters in Van Tieghem's Traité de Botanique (Paris, 1891) may be consulted, as should be Pfeffer's Pflanzenphysiologie, and Frank's Lehrbuch der Botanik, Band i. (Leip. 1892).
Vegetarianism is popularly defined as the practice of eating foods obtained from the vegetable kingdom exclusively, and abstaining from all foods obtained from the animal kingdom. This is an approximately correct definition of what was meant by the word when the Vegetarian Society was founded in 1847. It was not long, however, before it was discovered that there was as great a need of discrimination in the use of the various products of the vegetable kingdom as there was cause to abjure the use of flesh. Vegetables are not all equally wholesome, some are absolutely poisonous. There is a great difference between the stalks and leaves and the seeds of plants. Cooked and uncooked foods differ greatly. The study of food leads to the study of equally important laws. Eggs, milk, cheese, butter (animal products) differ from the carcasses of slaughtered animals. Many of the arguments against the use of flesh do not apply to the use of fish. These and similar questions arose, and so sects sprang up. Some ate fish; some were called Vems (eating Vegetables, Eggs, and Milk); some were strict vegetarians or Vego (excluding all animal products); some took up the Edenic diet (excluding all cooked food); some, though using cooked food, excluded cereals; some adopted a theory of a specific vitality in uncooked fruits, grains, nuts, and pulses, specifically adapted to the development of man on his four planes of body, mind, soul, and spirit. Out of this clash of conflicting ideas there gradually grew up in the London Society a symmetrical theory which has widened the basis of vegetarianism (Neo-vegetarianism). It holds that nature is governed by universal law; that health, happiness, and life are normal, and disease, misery, death abnormal; that man has power to regain the normal by obedience to the laws of nature (the abnormal being the result of inherited or personal disobedience); and that vegetarianism is coincident with obedience to these laws.
In strictness, therefore, no one is yet entitled to the name of vegetarian, but since it is believed that the eating of flesh is antagonistic to progress on so many grounds, and that reform in food is the basis of all individual reform, the renunciation of flesh in whole or in part is made the test question of vegetarianism, leaving the positive question of what articles in the vegetable kingdom constitute man's proper food to be one of the branches of further study. In support of abstinence from flesh the following ten claims are made: I. Physiology proves man to be closely akin to the frugivorous ape and essentially different from the carnivora, herbivora, or omnivora. But it is objected that man has canine teeth, and his intestines of mid length between the herbivora and carnivora show that he should partake of the food of both. To this the reply is that canine teeth are found in apes also; and the mean length of his intestines proves rather that he is fit for neither than for both forms of food, for grasses need long intestines which he has not got, and flesh needs to be passed away very rapidly through intestines shorter than man's, to prevent putrefaction. Physiologically and anatomically man can scarcely be differentiated from the higher apes. II. Embryology shows that to be a permanent differentiation of type which comparative anatomy shows to be actually different: the typical herbivora being placentally non-deciduate, the carnivora zonary deciduate, and apes and men discoidally deciduate. III. Chemistry proves that all elements for perfect nutrition in assimilable forms are found in a proper vegetarian dietary. It is objected that (a) an enormous bulk must be consumed to get the requisite nutriment; (b) flesh is food ready prepared by another animal, and so requires less vital strain to assimilate it than vegetable food. It is replied that (a) increased bulk is not necessary—e.g. of flesh alone it would be necessary to eat 96 oz. per day to get sufficient carbon; of bread alone, 64 oz. to get enough proteins; of mixed flesh and bread, 42.3 oz. (34.5 oz. bread and 7.8 oz. beef) would suffice; similarly 34.6 oz. of a mixture of 9.5 oz. lentils and 25.1 oz. of bread, or 24.1 oz. of almonds and raisins, would give the same result; (b) animal cells are not absorbed as cells, but must be broken down and converted to chyle before absorption, so that assimilableness varies as much between different sorts of flesh as between flesh and non-flesh foods. Thus pork takes, raw, 3 hours; well roasted, 5½ hours; beans, 3½ hours; beef, raw, 2 hours; well boiled, 2¾ to 3 hours; well roasted, 3¼ to 4 hours; rice, boiled, 1 hour; fresh-baked bread, 3½ hours to digest. IV. Flesh as sold is diseased to a very high percentage by diseases communicable to man, which the purchaser is quite unable to detect. V. Higher instincts revolt against taking life, and the repression of these higher instincts prevents evolution of the higher man; while the relegation of the function of slaughtering to others is the unjust perpetuation of a degraded class to be an ever-constant source of danger to the stability of the morality of the community for the sake of supplying the luxurious demands of an artificial appetite. The objection made is that butchers are often kind and gentle, and that other occupations produce types of men as low as slaughtermen. VI. Unnecessary destruction of life is immoral. Objections are made—(a) that it begs the question to say that flesh food is unnecessary; (b) the land would be overrun if the animals were allowed to breed unhindered; (c) is it not more merciful to breed, tend, feed, and painlessly kill, than allow the animals to battle for existence and then die of hunger or fall a prey to the carnivora? The reply to these difficulties is—(a) physiology and chemistry prove flesh to be unnecessary; (b) the balance of nature prevents overrunning—e.g. is any new land when discovered overrun with animals, although they have had countless ages in which to propagate unhindered by man's interference? (c) the problem presented is hypothetical, for in reality the cruelties inflicted by man on the animal world are appalling, and for these cruelties the habit of flesh-eating is to a great extent responsible. VII. It is economically superior—for 1.281 lb. of oatmeal at a cost of 3d. will supply power to raise 140 lb. to the height of 10,000 feet, while of beef fat it would take 555 lb. at a cost of 4½d., or of beef lean 3.532 lb. at a cost of 3s. 6½d. On the other hand it is said that all were to adopt it it would simply mean an all round depreciation of wages by a reduction of the subsistence minimum. VIII. Physical strength increases, as proved by the usually superior physique of peasants to that of middle classes; Arab porters, Indian runners, and Scotch and Irish peasants are especially pointed out as physically above the average, while the frugivorous gorilla is noted for enormous strength. But it is objected that (a) men who are so fed improve when flesh is added to their diet; (b) many Hindus are puny and staminally weak. It is replied that (a) they appear to improve because flesh is a stimulant, and so transforms latent energy into manifest force; (b) it is not claimed that a diet chiefly composed of rice is the best, while the fact that many fine races do live without flesh proves that under such conditions at any rate it is not a necessity; and immediately the non-necessity is admitted the whole weight of the moral argument enters the scale against the habit. IX. Whereas 12 acres of land, if used for the rearing of cattle for slaughter, will maintain one man feeding on the flesh produced, the same area under wheat will maintain twenty-three, and on a mixed crop of fruit, pulse, grain, and vegetables a still higher number. The objection here is that much land is suitable for nothing else but, e.g., sheep runs. By the vegetarian this may be granted, but the assertion does not apply to the thousands of acres of valuable horticultural land now under grass. X. It is claimed that horticulture, and especially petite culture, would employ an enormously greater amount of labour than does stock-raising, and so tend to afford a counter current to the present toward drift and to congested labour centres. Vegetarianism has spread so extensively that in addition to its pledged adherents it has a still greater number who practise abstinence from flesh to a greater or lesser degree in obedience to medical advice, for the curing or relieving of disorders of digestion, gout, and rheumatism. The Catholic Church enjoins abstinence from flesh during Lent, and on some other days during the year. Many religious orders (e.g. the
Trappists) abstain wholly from flesh. Brahmins also abstain from flesh and eggs. The Vegetarian Federal Union was formed in 1889. Its offices are at the Memorial Hall, Farrington Street, E.C. To it are affiliated the Vegetarian Society, the London Vegetarian Society, eighteen other English societies, and those of America, Germany, and Australia. An international congress was held at Cologne in 1889, in London in 1890. There are now vegetarian restaurants in many large towns.
The chief publications are The Vegetarian (weekly, Lond.); The Vegetarian Messenger (monthly, Manchester); Food, Home, and Garden (monthly, Phila.); Der Vegetarier (bi-monthly, Berl.); Die Neue Heilkunst (bi-monthly, Berl.). See John Smith, Fruits and Farinaea; Alcott, Vegetable Diet; Professor Francis W. Newman, Essays on Diet; Howard Williams, Ethics of Diet—all obtainable from the Veg. Federal Union; also Springer, Wegweiser durch die Veg. Litteratur (2d ed. 1880).