Coral, a term loosely applied to any animal in the class Cœlenterata which forms a hard skeleton. The tubular organ-pipe coral, the noble or red coral of commerce, and the reef-building madrepore corals, are familiar examples. Presuming an acquaintance with the general features of the class, as summed up in the article CŒLENTERATA, we may notice at the outset the fact that the formation of hard supporting structures is exhibited in very varied degrees, in manifold styles of architecture, and in widely separated forms.
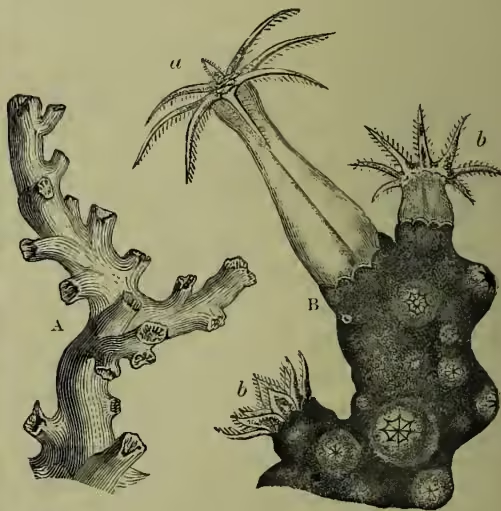
Different Kinds of Coral.—(1) Among the Hydrozoa the skeletal investment of the polyp types, when present, is usually horny. In one subdivision, however, the supporting framework is limy, and to these forms—Millepores (q.v.) and Stylasterids—the title Hydrocorallina is fitly applied. (2) It is, however, in the other sub-class types, when present, is usually horny. In one subdivision, however, the supporting framework is limy, and to these forms—Millepores (q.v.) and Stylasterids—the title Hydrocorallina is fitly applied. (2) It is, however, in the other sub-class of Cœlenterates—the Actinozoa—that the formation of coral structure becomes emphasised and perfected. In many simple and in most colonial forms, a skeleton is formed, as isolated spicules or continuous. When continuous it may be horny, or limy with an organic basis, or to all appearance entirely limy. (a) Among the Alcyonaria, of which Alcyonium (q.v.) is a convenient type, Tubipora, with the spicules united to form tubes and crossing platforms; Corallium, with a branched axis of fused spicules; Isis, with a jointed axis alternately limy and horny; Heliopora, with limy cups enclosing the individual animals, and traversed as they grow by successive horizontal floors at slight intervals, are examples of different forms of coral occurring in distinct groups. (b) Among the Zoantharia, of which Actinia or any sea-anemone is a familiar example, the Actinaria have no skeleton, and therefore include no coral forms; the Antipatharia have a varied non-calcareous supporting skeleton, and include some forms known as 'black corals'; the third subdivision, the Madreporaria, always have a continuous limy skeleton, and include the best-developed corals. Most madrepores are colonial, and the common connecting skeleton is either solid (apart from mere gaps) or perforated by canals which establish community of life between the individual members. Hence it is usual to distinguish two sections of Madreporas as Aporosa and Perforata. (3) The earliest corals of Palæozoic strata—the Rugosa or Tetracoralla—are distinguished by four primary partitions.
General Structure.—In discussing further the general structure, it will be convenient to leave the widely separated Hydrocorallina out of account. The most general fact that can be stated at the outset is, that with the possible exception of the sea-pens, the cells which form the skeleton are at least originally epidermic. Free spicules, connected lattice works, solid cups, illustrate various degrees of lime deposition; while supporting axes, ensheathing tubes, and what look like tolerably complete casts of the entire animal, are the main types of coral architecture. In detail, however, those main types vary widely, and in the cup-corals especially, the structure is often very complex. In a simple madrepore coral, representing a single individual, the following structures are at once obvious. The main cup-like mass, known as the theca, is usually attached by its base to some foreign object. A terminal depression—the calyx—contains the animal, the soft parts of which extend more or less over the lips. Within the cup, when the animal has died away, radial partitions are seen, forming various systems of septa. Some of these often meet in the middle of the cup to form a central pillar or columnella. When the coral is colonial, as is generally the case, complications arise by the fusion of individuals. The deeper parts of growing corals are sometimes separated off by cross partitions (tabulae), but this is rare in the Zoantharia. In large masses of madrepore certain portions of the skeleton are often left behind by the animals, and being exposed to the water, become corroded and altered. In regard to the development of corals, it is hardly possible yet to make any useful general statements. For the animals themselves, it must be enough here to refer to the articles on ALCYONIUM and ANEMONE, which, though not themselves 'corals,' are good examples of the two sub-classes to which most corals belong.
General Life.—Corals are predominantly passive forms, and are much modified by their environment (currents, &c.); the great majority are permanently fixed, except in embryonic life. Flabellum may be noted as an instance of a coral-cup which becomes free as an adult. The limy material which forms the skeleton seems, in many cases at any rate, to be derived not from the water, but from the minute animals which constitute the food. The great majority are colonial, yet cases of division of labour are very rare. Many corals are richly coloured, but the meaning of the numerous pigments is still imperfectly known. Masses of corals furnish browsing ground for crowds of animals, not a few of which intrude into the colonies, while others have become established parasites. Some corals also include parasitic Algae (see SYMBIOSIS). Semper describes some very interesting cases of the constant association of worms and corals, and notes how the guest has permanently modified the host (see COMMENSALISM). It is interesting to notice, apart from actual modification of form, how various worm-organisms act upon the 'sea-meadows of coral' much as earthworms do upon land-meadows (see Huxley's Invertebrates, p. 171).
In most corals the sexes are separate, and even the colonies may be entirely male or female. In Corallium, hermaphrodites may occur, or different branches be of either sex. As is common in sessile passive organisms, asexual or vegetative reproduction is a marked feature. Semper considers the budding of the Fungia colony as illustrating 'alternation of generations.'
Distribution.—Of the madrepore corals, the 'solitary' forms, which never form great masses, are for the most part widely distributed deep-sea animals, while the reef-builders are as markedly dwellers in shallow water, from the low-tide mark to about 20 fathoms, and are limited to warm waters. They are not found on the western coasts of Africa and America. In the waters of the Central Pacific reef-corals are found in greatest profusion and variety; but those of the Red Sea, Indian Ocean, and East Indies are not far behind. Over eighty fossil genera of the Palæozoic Rugosa are known, and of the other Actinozoan corals, about 1800 fossil, and as many living species. Of these, the vast majority are madrepores, and only a small number Alcyonarian. Both sets begin in Silurian times, but were for long outnumbered by the Rugosa. Most existing genera date only from the Tertiary period. The 'areas of past distribution of coral-reefs in no way correspond with those distinctive of the modern seas.' See Heilprin, Distribution of Animals (1887).

Important Forms.—The most important corals are the reef-builders (Porites, &c.). The delicate colour, exquisite architecture, and labyrinthine complexity make the limy skeletons beautiful objects. Yet more beautiful, however, is the wealth of colour often exhibited by the living forms. Caryophyllia is a simple and solitary cup-coral found on the south coast of England. Bathyactis and Deltocyathus are almost world-wide. Antipathes—the black coral—has a valuable hard axis like ebony. Fungia is worth noting on account of its flat mushroom-like shape and Semper's observation (see above) in regard to alternation of generations; among Alcyonarian forms may be noted the blue coral (Heliopora cærulea) of many Pacific coral-reefs, the horny and beautifully coloured sea-fan (Gorgonia flabellum), and the familiar organ-pipe coral (Tubipora musica).
More important, however, in practical interest is the red coral (Corallium rubrum) of commerce. In this coral the limy spicules are fused to form the familiar solid axis. The most important banks are those of Algiers and Tunis, and off the Sardinian, Sicilian, and Balcaric islands. Great quantities are fished near Naples, and at Sciacca in Sicily. The coral is roughly dredged, freed from its rind, assorted according to colour, and manufactured in Naples, Leghorn, Genoa, &c. Pale rose-coloured coral, not spoilt by the boring of worms and other intruders, has fetched £12, 10s. per ounce. The industry is a very important one, though in recent years it has seriously declined. The most important markets are in Asia, Africa, parts of Russia, Japan, and South America.
CORAL ISLANDS is the name given to certain low islets which are composed for the most part of the calcareous skeletons of various kinds of corals. An atoll or typical coral island consists of a somewhat ring-shaped reef inclosing a lagoon. Such reefs vary in size from less than a mile up to 90 miles long and may be 10 miles wide—the breadth of the annular reef being on an average about a quarter of a mile. Barrier-reefs, which have a similar breadth, are found surrounding islands or bordering mainland, from which they are separated by a navigable channel. Fringing-reefs, on the other hand, extend outwards from the shore, with no separating lagoon-channel. In the case of atolls and barrier-reefs there is generally a navigable passage through the reef into the lagoon or lagoon-channel, which is kept open by the scour of the tide. It is upon atolls or barrier-reefs that islands occur. Only very rarely, however, are these islands co-extensive with the reef—generally they appear as longer or shorter belts, forming a series of islets straggling along its surface. As reef-building corals do not flourish in water having a lower temperature than 68° F., they are necessarily restricted to tropical and subtropical seas—from many regions of which, however, they are excluded by the presence of cold currents coming from extra-tropical latitudes, by the muddy character of the water opposite the mouths of large rivers, and by other causes.
The rock of a coral-reef is a white limestone composed chiefly of masses of corals, coral debris, and sand, together with the hard parts of molluscs, echinoderms, bryozoans, and calcareous algæ. Not infrequently this rock has been changed by the chemical action of percolating water into a compact or crystalline mass, which has lost all, or nearly all, trace of its organic structure. Chamisso (q.v., known also as a poet), the naturalist who accompanied Kotzebue the Russian navigator in his voyage of discovery into the South Sea in 1815–18, appears to have been the first to show how such reefs are converted into dry land. He describes how the polyps cease to grow when they reach the surface of the sea, and how the reef, exposed at low-tide, by and by becomes disintegrated, while broken coral debris is heaped up by the action of the breakers so as eventually to form a high bank which can only be covered during spring-tides. This bank of coral blocks and debris solidifies in time, and remains exposed to the fierce heat of the sun, so that the mass shrinks and cracks, and flakes are detached from it, and raised one upon another by each returning tide. ‘The always active surf throws blocks of coral (frequently of a fathom in length and 3 or 4 feet thick) and shells of marine animals between and upon the foundation-stones; after this the calcareous sand lies undisturbed, and offers to the seeds of trees and plants cast upon it by the waves a soil upon which they rapidly grow to overshadow its dazzling white surface. Entire trunks of trees, which are carried by the rivers from other countries and islands, find here at length a resting-place after their long wanderings: with these come small animals, such as lizards and insects, as the first inhabitants. Even before the trees form a wood, the real sea-birds nestle here; stray land-birds take refuge in the bushes; and at a much later period, when the work has been long since completed, man also appears, builds his hut on the fruitful soil formed by the corruption of the leaves of the trees, and calls himself lord and proprietor of this new creation.’ According to Professor L. Agassiz, the large blocks torn off by the breakers are loosened not by shrinkage under the sun’s heat, as Chamisso supposed, but by the innumerable perforations of various boring molluscs.
The origin of coral-reefs is a question which has at various times given rise to discussion. According to Chamisso, the corals commenced to grow on shoals in the sea—on the tops of submarine mountains in short. The circular form of the atoll and its basin-shaped lagoon he believed to be due to the natural growth of the coral. The most vigorous growth took place upon the outward edges of the reef, where the largest and most massive corals flourished. In the interior the coral growth was hindered by the accumulation of shells, coral debris, and sand swept forward by the waves. The reef, therefore, as it approached the surface would gradually assume a basin-shape—the outer edges, or peripheral and actively growing portion, forming the reef—the interior part, where growth was retarded or prevented by sediment, &c., forming the lagoon. Other writers attempted to account for the annular form of atolls by supposing that the reefs had grown upwards from the crests of submarine volcanic craters. The not infrequent occurrence of volcanoes and volcanic islands in the coral regions of the Pacific Ocean was thought to favour this supposition, but the enormous diameter of many atolls seemed to preclude the possibility that these larger atolls, at all events, could be founded on the lips of submarine craters. And when at length it was discovered that reef-building corals flourish only in comparatively shallow water, this hypothesis was rejected, since it was extremely unlikely that so many submarine volcanic mountains should rear their summits just to the limits within which the corals could begin their superstructures.
Darwin, during the famous voyage of the Beagle, was naturally fascinated by the problem of the origin of these remarkable reefs, and while he admitted that certain atolls might very likely have been formed in the manner described by Chamisso, yet he could not believe it possible that a broad mountain-summits lay buried at the depth of a few fathoms beneath every atoll, and nevertheless that throughout the immense areas occupied by many of those reefs not one point of rock should project above the surface. The theory of subsidence which he subsequently advanced seemed to account for all the phenomena, and until recently it commanded nearly universal assent. According to Darwin, each atoll has passed through the successive stages of fringing-reef and barrier-reef. Since reef-building corals do not thrive at greater depths than 100 feet or thereabout, it is evident that the foundations of a coral reef could not have been laid in deeper water. And as such moderate depths occur only round islands and off the shores of continents, the reef-builders would begin their work by forming at first a fringing-reef. Slow subsidence of the sea-bottom is supposed to have supervened; but while the foundations were being carried down, the corals continued to grow upwards, the rising of the reef keeping pace with the sinking of the sea-bottom. Thus by-and-by the fringing-reef is converted into a barrier-reef. We have now only to suppose that the movement of subsidence and the labours of the corals continue until the reef-encircled island disappears below the waves, and a complete atoll will be the final result. Darwin's views were illustrated and strongly supported by Professor J. D. Dana, who accompanied the Wilkes exploring expedition round the world in 1838-42, while M. M. Couthouy and Beete Jukes likewise upheld the hypothesis of subsidence. The researches of Professors L. Agassiz and Le Conte amongst the reefs of Florida have shown that Darwin's views are there inapplicable. They attribute the formation of the southern portion of Florida, which consists of a series of concentric barrier-reefs, to the natural growth of the coral alone aided by the mechanical action of the sea—a view which, stated generally, is the same as that held by Chamisso. Professor Karl Semper likewise, after examining the reefs of the Pelew Islands, came to the conclusion that the form of these reefs had been determined by the natural growth of the corals, modified by local conditions, which is just Chamisso's explanation. But Darwin had already expressly stated that some reefs might have originated in the manner suggested by Chamisso; and geologists apparently recognised that the conditions of the great Florida reef were hardly analogous to the isolated atolls which rise from profound depths in the open ocean.
In 1880, however, Dr (afterwards Sir) John Murray of the Challenger expedition, published another explanation which has given rise to considerable discussion. He returns to the old views of Chamisso, maintaining that reefs have grown up from the tops of submerged and partially submerged banks and mountains. To the objection that it is hard to believe that so many banks and cones should occur just at the proper depth from the surface, Dr Murray replies that it is not necessary to suppose that all the submarine eminences were of equal or nearly equal height. Some may have risen at first above the level of the sea, and subsequently have been reduced by the waves and breakers to the condition of shoals; while others that did not reach to the limits within which reef-builders live may have been brought up to that zone by the accumulation upon them of the hard parts of pelagic organisms. For these forms of life flourish in extraordinary numbers in the surface-waters of the tropics where reefs abound, and as they die, their hard parts falling to the bottom will accumulate there along with the exuviae of creatures living at such depths, until they come to form considerable deposits. This levelling-up of submarine banks to the zone within which coral-builders thrive is the most marked feature in Dr Murray's hypothesis. His further account of the mode in which atolls and barrier-reefs have been formed is a more particular and scientific development of Chamisso's explanation. The growth of a reef is regulated by the food-supply of the corals; those growing on the external parts of the reef are most favourably placed in this respect, consequently they thrive best, and have the advantage over those that are growing in the centre of the reef; and this difference in the rate of growth of the outer and inner areas will be intensified as the reef approaches the surface. Eventually the corals of the interior die, and the dead coral rock is gradually removed by the solvent action of the sea-water which contains carbonic acid. In this manner the reef assumes a basin-shaped form. Fringing-reefs, according to Dr Murray, are converted into barrier-reefs by the simple outward growth of the coral upon a talus of its own debris, forced off from the edge of the reef by the breakers. The lagoon-channel that eventually separates the reef from the land is formed in the same way as the lagoon of an atoll—namely, by the dissolution and removal of dead coral rock. Professor A. Agassiz published in 1882 an account of his detailed examination of the Tortugas and the Florida reefs, in which he shows how the bottom of the sea in those regions has been gradually raised by the accumulation of the exuviae and skeletons of massive organisms which flourish in prodigious numbers on the sea-floor, forming the submarine plateaus known as the Florida, Yucatan, and San Pedro banks. He thinks it is by the gradual upward growth of such accumulations that the sea-bottom was eventually raised to the zone in which reef-builders thrive. He thus arrived independently at the same general conclusion as Dr Murray.
As recently as 1887 Dr Guppy (q.v.) has given the result of his investigations in the Solomon Islands, and has vigorously supported the hypothesis advocated by Murray. He shows that in those islands we have elevated reefs which exhibit the very structure which we should expect to find if the new view of coral-reef structure is correct. He found some of those islands largely composed of earthy calcareous deposits having the same character and origin as the various volcanic muds and organic oozes which the Challenger expedition dredged at depths varying from 100 to 2000 fathoms. These deposits reached to heights of over 1000 feet, and in some cases denudation had exposed an underlying nucleus of volcanic rock. The earthy calcareous beds were covered with a comparatively thin crust of true coral rock. The phenomena described by Dr Guppy thus lead to the belief that many other atolls may have a similar structure. Dr Guppy thinks that the various agencies insisted upon by Semper, Murray, and A. Agassiz—such as currents and tidal scour, solution and the distribution of food-supply, the repressive influence of sediment and the action of the breakers—have each played a part in determining the form of coral reefs. To the objection that some lagoons are deeper than the zone in which reef-corals live, Dr Guppy replies that where the submarine slope is moderate, the water clear, and breakers of no great size, reef-corals may be found flourishing at depths of 50 and even 60 fathoms. Whether the explanation advocated by Murray and Guppy is destined to lead to the total collapse of Darwin's hypothesis, is likely to remain for some time an unsettled question—some writers holding that while Murray's view may be the true explanation of the origin of many atolls and barrier-reefs, it does not forbid the possibility or, as others would say, the probability that many other reefs may really have been formed, as Darwin supposed, during slow subsidence of the bottom.
See CÆLENTERATES, ANEMONE, ALCYONIUM; textbooks of Zoology—Huxley, Claus, &c.; Darwin, Dana, Murray, Guppy, &c. on Coral-reefs, and Semper's Animal Life; Moseley and Quelch Challenger Rep., vols. ii. and xvi.; for skeleton, Von Koch, Morphologisches Jahrbuch (1879-86); for red coral, Lacaze-Duthiers, Histoire Naturelle du Corail (1864); for fossil forms, Zittel's Palæontologie; systematic works—Dana, Zoophytes (1848-49); Milne-Edwards and Haime, Hist. Nat. des Coralliaires (1857-60); Koren and Danielssen, Nye Alcyonider, Gorgonider og Pennatulider (1883); Koch, Monograph on Gorgonida (Naples Station, 1887).