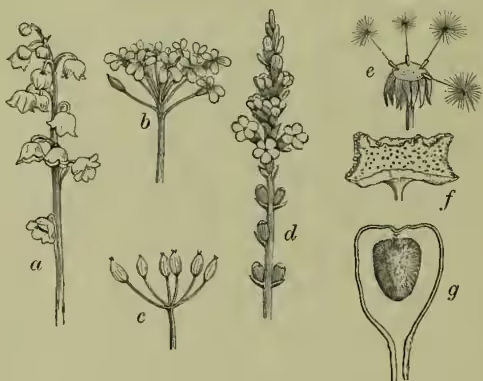
Inflorescence. This term is applied by botanists in a concrete and special, as well as in an abstract and general sense—i.e. first to any single group or natural aggregate of flowers arising upon a common main axis, and secondly to the various modes or principles of floral arrangement themselves. Despite that endless superficial diversity upon which the characteristic aspect of different species and larger groups so much depends, these apparently indefinite variations may readily be reduced to a small number of easily intelligible types. For, while the earlier botanists naturally tended to develop a nomenclature corresponding to the multiplicity of outward forms which inflorescences acquire, the progress of research has simplified this by centering attention upon the few and simple modes of branching by which they arise. We naturally set out with any plant of which the axis continues to grow indefinitely, but of which a number of secondary axes arising in the axils of the leaves are developed as flowers. When the pedicels of these flowers tend to reach a moderately equal length the inflorescence is known as a raceme (fig. 1, a); or when the process of floral development arrests them, so that the flowers are practically sessile, we have a spike (fig. 1, d). The growing point of most racemes and spikes, however, tends to be checked by the reproductive stress, and the axis thus frequently ends, or rather seems to end, in a terminal flower. Good examples of this essentially racemose or spicate type are furnished by many Liliaceæ, Scrophulariaceæ, &c.— e.g. Tritoma, foxglove, mullein, &c. Even such a curious inflorescence as that of the pine-apple may now easily be interpreted as a greatly condensed spike of fruits, crowned by its leafy growing point.
The shortening of the main axis of a raceme may take place after ordinary development has begun, so that the upper internodes are much less developed than their predecessors. The pedicels of the younger flowers naturally share the same arrest of development, and thus it is that the comparatively long pedicels of the lower flowers place them on much the same level as the higher ones, and even as the summit of the axis. This variety is known as the corymb (fig. 1, b), so familiar in the candy-tuft. When all the internodes are so shortened that the pedicels arise from practically the same level, we have the umbel (fig. 1, c), so characteristic of the Umbelliferae.
Suppose this vegetative arrestment and floral precocity to be continued still further, internodes and pedicels alike become arrested, and the result is a crowded cone or excessively shortened spike of sessile flowers. By continuing the same process which gave us the corymb, the cone necessarily tends to appear more and more depressed through the more rapid upgrowth of its lower portions; thus we have that characteristically expanded axis, so compactly set with florets as to resemble at first sight a single flower, familiarly known as the head or capitulum of the Composites (fig. 1, e). The spiral arrangement of the florets so obvious in a sunflower is simply that of the depressed cone, which we may again draw out in imagination into the corymb, the spike, or the raceme, with which it is in principle identical.
A capitulum fundamentally similar may, however, be evolved in a slightly different way, by the more or less complete arrestment of the secondary axes of an umbel. Hence it is that a few umbelliferous plants, like Astrantia, or still better the sea-holly (Eryngium), &c., come to present that appearance which so often induces the beginner to confuse them with Composites.
But, since it is manifest that the same embryonic shortening may occur in any type of inflorescence whatsoever, it becomes evident that we must reserve the term capitulum for the type of inflorescence presented by the Composite or Scabious, leaving the various superficially resembling forms or pseudo-capitula, as of sea-holly already referred to, or those of sea-pink, of bergamot, &c., to be separately analysed according to their true origin.
Returning, then, to the capitulum proper, we must continue to keep clearly in view that conception of vegetative growth (as reaching its maximum rate only at some distance behind the growing point) which may be actually verified by measurements of any growing shoot or root. The conical axis thus not only tends to broaden and flatten, but its lower portion must at length overtake the apex, and a perfectly flat receptacle, as in some species of Dorstenia, results. The margins next outgrow the apex, and the cone is now becoming a shallow saucer (other species of Dorstenia, fig. 1, f). The saucer next becomes a cup, or even flask; and the remarkable hollow inflorescence of the fig (fig. 1, g) is thus seen to be morphologically akin to the capitulum, and through this by the corymb even to the original raceme itself.
The study of vegetative branching (see BRANCH) has, however, shown us that we may have to do with compound or sympodial axes as well as simple or monopodial ones. That is to say, in our primary axis the growing point may perish, leaving, however, of course, all the more opportunity for the development of the secondary axes latent in its lateral buds. This disappearance of the primary growing point, having once set in, soon works back, until we have it occurring immediately after the development of the first lateral bud. This then readily takes its place for practical purposes, just as a larch or pine which has lost its top renews it by the upgrowth of a branch. But the new axis dies in turn after giving birth to its successor, and so on; thus the false axis or sympode is formed. Inflorescences of this type are known as cymes. The simplest in principle is that of the Day-lily (q.v.). It is commonly known as the helicoid cyme (fig. 2, h), since the origin of the new axes winds on in the same spiral order as that of the leaves upon the primary axis itself. The distinction from a raceme is, however, easily made when we notice that the so-called bracts are not really bracts at all, but are more or less opposite to the flowers; being really only the axillant leaves of the next axis, which bears its flower only after producing a leaf with the bud of its successor.
But in other cases the spiral may change its direction with each new axis, and the false axis thus assumes a very different appearance, that of the scorpioid cyme (fig. 2, i), of which the classical example is furnished by the Boraginaceæ (Goebel, however, regards these as unilateral racemes, and offers Tradescantia, Echeveria, &c. as more real types). This reversal of the spiral has been prettily verified by noting how in the scorpioid cyme of the
Rock-rose (Helianthemum) the spiral of the calyx runs in an opposite direction in each successive flower.
So far we have been dealing with cymose inflorescences as arising in plants with alternate leaves: in opposite-leaved plants—e.g. Caryophyllaceæ and Begonia (q.v.)—the resultant form is necessarily very different. Let the growing point terminate in a flower as before; but since each of the two leaves immediately below is in an equally favourable condition, both as regards radiation and alimient, we have two secondary axes instead of one. Hence, instead of one secondary axis continuing in the line of the primary one, we have necessarily two of equal strength and divergent at an equal angle. The main axis thus at first sight
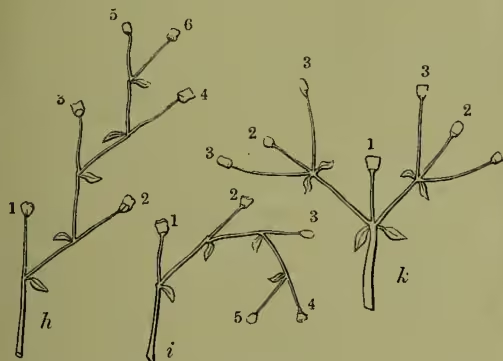
Diagrammatic representation of h, helicoid cyme; i, scorpioid cyme; k, dichasium. seems to have forked, especially when the terminal flower disappears, just as in the false dichotomy so apparent in the branches of the lilac or mistletoe; and hence the old name of dichotomous cyme, which, however, it is evidently necessary to correct, as dichasium (fig. 2, k), biparous cyme, or the like.
This inflorescence may undergo shortening, or in more physiological language remain more or less embryonic, as in most Labiatae (which, however, present all gradations, from the fully-developed cymes of Hyssopus, through the 'false whorls' or 'verticillasters' of the majority of genera, to the terminal pseudo-capitulum of Bergamot). The apparent umbel of geraniums and the pseudo-capitulum of the sea-pink have also this origin.
Not only modifications of these leading types, but various combinations, simple, compound, and complex, and in all degrees of reduction or exuberance, may also arise; the question of separating all the preceding types of inflorescence as radial from a small residuum as dorsi-ventral also presents itself. See Goebel's Outlines of Classification and Van Tieghem's Traité de Botanique.