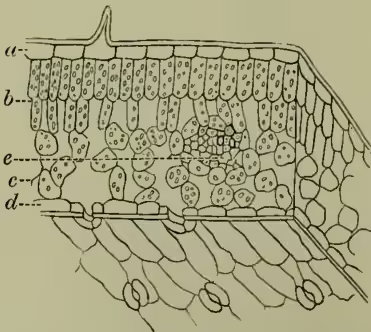
Leaf. Leaves are lateral organs developed from the stem or Axis (q.v.) of the plant below its growing point. They never bear flowers, and after reaching their full development they retain their form and size unchanged until death, after which they are removed from the stem either by gradual decay (most monocotyledons) or by breaking off at a distinct articulation (most dicotyledons). They normally consist of two main parts, a stalk or petiole, and a blade or lamina, the latter being usually flattened and expanded. They may also possess lateral appendages or stipules at the base of the petiole. Physiologically considered, they are of the highest importance, as can best be understood after examination of their minute anatomy. The petiole resembles a stem in structure; the blade, however, is distinguished by the great development of cellular tissue, through which the fibro-vascular bundles pursue a course usually similar to that which they possess in the stem, thus exhibiting the parallel and reticulated venation so characteristic of Monocotyledons (q.v.) and Dicotyledons (q.v.) respectively. Taking dermis, which exposes the subjacent (2) cellular ground tissue or parenchyma, which is dark green on the upper, and paler because of looser texture on the lower side, while (3) the fibro-vascular bundles can readily be prepared as a skeleton by scraping, or better by maceration. A thin transverse section placed under the microscope shows, proceeding from above downwards, (1) the upper epidermis, a continuous layer of empty cells, with walls often considerably thickened, especially on the upper surface, to form the so-called cuticle; (2) the parenchyma, which contains the fibro-vascular bundles, and which is readily distinguishable into two chief layers. Above is the so-called 'palisade parenchyma,' in which the cells are elongated vertically and placed close together like the posts of a palisade, and below this lies the loose parenchyma of the middle and lower surface of the leaf, in which the cells are polyhedral and loosely arranged, leaving irregular air-passages. Finally we have the lower epidermis, in which numerous openings, the Stomata (q.v.), place the spaces in the parenchyma in continuity with the external atmosphere.
The essential function of leaves resides in their chlorophyll-containing parenchyma, in which, in presence of light, carbonic acid () is decomposed with evolution of oxygen into the atmosphere and formation of starch (see CHLOROPHYLL, VEGETABLE PHYSIOLOGY). This process has, of course, nothing to do with the function of Respiration (q.v.)—oxidation of protoplasm with formation of carbonic acid which is going on constantly during life in all the tissues of plants as well as of animals; it is a compensatory process whereby the green plant is enabled to repair its respiratory losses of matter and energy, and provide for its continued growth by the fixing of new matter from the atmosphere and new energy from the sun. The newly-formed starch is first distinguishable in the form of granules, which are visible in the substance of the chlorophyll grains, is then digested into a soluble body, probably glucose, and carried off by the descending Sap (q.v.), to be either directly assimilated to form new protoplasm, or to be reconverted into starch and stored for future use.
The functions of the blade of the leaf are shared to some extent by the petiole, by the green cellular envelope of the stem and branches (which thus not unfrequently come to replace the leaves altogether, good respective instances being furnished by cactuses and acacias), and often by the calyx and ovaries; in short, every part of the plant exposed to light tends to utilise it by producing chlorophyll, excepting only those parts of the flower where, in current phrase, more conspicuous colouring matters are required for the attraction of insects.
The forms of leaves are greatly varied, often obviously in adaptation to the habit of the plant, large and free-growing plants which obtain unobstructed light most frequently bearing simple or slightly lobed leaves, while the smaller vegetation generally produces leaves either long, simple, and narrow (e.g. grasses), or highly compound, with small leaflets (e.g. ferns), so as to seize as many as possible of the broken sunbeams which have not been intercepted by the loftier plants, while casting as little shadow as possible upon each other. Again, the leaves of aquatic plants, if floating, are simple and largely expanded, so as to maintain their position and obtain the maximum of light (e.g. water-lily and pond-weed), but if submerged are usually dissected into filiform segments (water-primrose), so as to allow the water to flow unobstructed, and thus constantly renew the supplies of carbonic acid. Again, where in one and the same plant the leaves on the lower and upper common examples of such leaves, it is easy to make out all the principal tissues (see BARK); (1) thus, by tearing the leaf obliquely, we can remove shreds of dry, colourless, transparent epi- portion of the axis are in different circumstances, their form is also varied, and we have the heterophyllous condition, which can be seen in many land-plants, but perhaps most conveniently in the water buttercup (Ranunculus aquatilis), which possesses both floating leaves which are simple, and submerged leaves which are highly dissected. So, too, plants which grow in dry and sandy situations, and obtain scanty supplies of water, either owing to drought or to too pervious soil, very frequently store their water in their leaves, which thus become succulent, and preserve it from the evaporative action of the sun by the aid of a thickened epidermis containing unusually few stomata.
Again, leaves may acquire entirely new functions, and have their form altered in correspondence with these. Where the plant is a climber the whole or part of the leaf may be modified into a tendril; where it is insectivorous it may be converted into a fly-trap (see INSECTIVOROUS PLANTS); or, as in the very highly specialised Nepenthes, we may have the base of the leaf of ordinary form and function, the middle twining as a tendril, and the tip hollowed and enlarged into a complicated pitcher.
Where protection is required, new modifications present themselves; if herbivorous mammals threaten the existence of the plant, the leaves or leaf-tips may become converted bodily into thorns, or covered with epidermic prickles. Bitter or acrid secretions, too, may develop, or stinging hairs be produced; while, if ants are to be guarded against, a hairy or glandular epidermis is the surest protection. Such at least are the interpretations commonly current (see DARWINIAN THEORY).
Such physiological considerations being grasped, comparatively little stress need be laid (save for purposes of specific description) upon the elaborate nomenclature of leaf forms with which botanical text-books are apt to be overburdened.
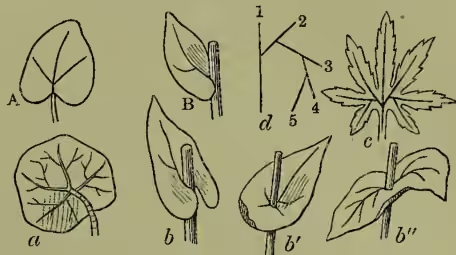
The parenchyma of the blade may be either in one continuous piece, when the leaf is said to be simple, or cut up into separate leaflets, when it is termed compound. Simple leaves may be conveniently reduced to three main forms, the circular, the elliptical, or the oval, according to the respective length and position of the longitudinal and the transverse diameter; the linear leaf being thus regarded as an elongated variety of the ellipti- cal, and so on. Innumerable variations in detail arise, however, according to the shape assumed by the apex, the margin, or the base of the leaf. The apex may be obtuse or acute, retuse or acuminate; the margin may be serrate or wavy, or parted into lobes so deep as to furnish transitions to the compound leaf; the base may be hastate or reniform, and so on. If the leaf base be prolonged beyond the insertion of the petiole and its lobes unite, we obtain the peltate condition familiar in the common Indian Cress (Tropæolum). If the petiole be absent, the leaf becomes sessile upon the axis; if its lobes are produced downwards, as in the reniform leaf, they clasp the axis, and the leaf is termed amplexicaul; if the lobes coalesce on the other side of the axis, it becomes perfoliate; and if they unite with those of a similar leaf arising on the opposite side, the pair are said to be connate.
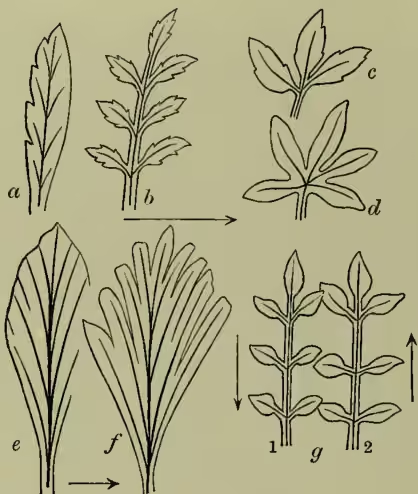
In compound leaves the leaflets may arise one from another on each side of a median lobe, as in the pedate leaf of Hellebore, or may radiate in palmate fashion from a common point—the end of the petiole, as in horse-chestnut; or, as is most frequent, they may be placed at intervals along the midrib, like the ribs of a feather, when we have the pinnate arrangement, of which the ash furnishes a familiar example. The simplest case of pinnate structure is where the lateral lobes or pinnæ are only two, as in the ternate leaf of clover: complex cases also are frequent, termed bipinnate, tripinnate, or decompound, according as secondary, tertiary, or even quaternary series of leaflets are developed.
How little morphological importance can be attached to these countless variations of form is well illustrated by the study of the development of the apparently similar 'pinnate' leaves of palms, dicotyledons, and ferns. In the palm the pinnate character is seen to be due to a mere tearing of a primarily simple leaf, by the midrib continuing to elongate after the parenchyma is developed; in dicotyledons the lobes develop separately, but sometimes from above downwards, and sometimes from below upwards; while in ferns the leaf is produced by a series of regular bifurcations of the growing point alternately to right and left, the first pinna being thus equivalent to all the rest of the leaf, and the apparent midrib a false axis, resulting from numerous separate dichotomies.
The comparative morphology of leaves is of the greatest interest. The essential conception, which floated before the eyes of Wolff and of Linnæus, was renewed by Goethe, and systematised by De Candolle, is that of a fundamental correspondence or serial homology among all the outgrowths from the sides of the axis—from the lowest and earliest, the seed leaves or cotyledons, upwards through the leaves proper to the bracts, and even thence through the parts arranged upon the floral axis—the sepals, composing the calyx, and the petals, forming the inner floral envelope or corolla, being still modifications of the leaf type, which we finally find most highly metamorphosed in the stamens and pistils (see FLOWER).
The transition from leaf to bract can be seen in any flowering plant, that from bracts to calyx may be conveniently studied in the mallow, that from sepals to petals in the cactus, that from petals to stamens in almost any garden rose (which indeed appears to have suggested the whole theory), and that from leaves to carpels in many monstrous flowers, especially the double cherry. Our consideration of the pinnate type of leaf-formation having shown that such apparent resemblances in adult anatomy are not necessarily real, it becomes necessary to test our theory by actual observation of the development of flowers. Embryology here furnishes an absolute confirmation—leaves and sepals, petals, stamens, and carpels, are all seen to develop as precisely similar processes of cellular tissue from the sides of the axis; and when the forms of leaves are fairly borne in mind, the apparent anomalies of flower structure become clear. Thus, the outer calyx (epicalyx) of a strawberry is readily seen to be composed of the united stipules of the sepaline leaves, the numerous stamens in five bunches of the St John's wort become resolved into a whorl of compound staminal leaves, and so on. While petals are obviously modified leaves, there is ground both developmental and analogical for regarding them, in some if not all cases, as barren stamens specialised to the attraction of insects; their relation to the leaf type becoming more remote (see STAMEN; and Grant Allen in Nature, July 1882).
The arrangement of leaves upon the axis (termed phyllotaxis) is always definite, and possesses a high degree of interest, although perhaps rather mathematical than morphological. An ascending spiral line may in all cases be traced round the axis through successive leaf bases, and these are found to occur at fixed distances, including a certain fraction of the circumference, most commonly , , , , or , although higher fractions of the same 'convergent series'—viz. , , &c.—or fractions of different series, and even transitions from one system to another, also not unfrequently occur, especially in such complex arrangements as the scales of fir cones. When leaves are opposite there are two primary generating spirals; when whorled there are three or more. The mode in which leaves are folded in the bud, termed prefoliation or vernation is of interest, since it is definite for each species. See Van Tieghem's, Sachs's, Vines's, and other text-books; also Lubbock's Flowers, Fruits, and Leaves ('Nature' series).