Seed. In the higher plants, which are called Phanerogams because of the conspicuousness of their flowers or reproductive organs, the egg-cell lies within an Ovule (q.v.), and after fertilisation grows into an embryo plant, with one or two primary leaves—all before separation from the parent plant. What is separated, to begin in favourable conditions a new and independent life, is a seed, which may be defined as a ripe ovule containing an embryo plant. So distinctive of the Phanerogams is this mode of reproduction by seeds, that they are often and conveniently called Spermatophytes or Seed-plants.
From the article OVULE the reader will understand how the seed is formed, how a mass of tissue—the nucellus—borne by the carpellary leaf contains a female spore or 'embryo-sac,' whose nucleus divides into a female nucleus or oosphere (which will develop into an embryo if fertilised) and a number of other nuclei of minor importance. The fertilised oosphere, within the embryo-sac, imbedded in the nucellus, and surrounded by the coats of the ovule, develops into an embryo plant, and the whole structure is called a seed.
Structure of the Seed considered in relation to the Growth and Germination of the Embryo.—The segmented egg-cell within the embryo-sac gives rise to the embryo and to a 'suspensor' which moors it. The figure on p. 300 shows somewhat diagrammatically the embryo moored to the top of the embryo-sac by the suspensor; the base of the embryo next the suspensor is the region from which the radicle or young root will spring; the depressed apex at the opposite pole of the embryo is the region from which the plumule or young stem will shoot forth; the two sides form the young cotyledons or seed-leaves.
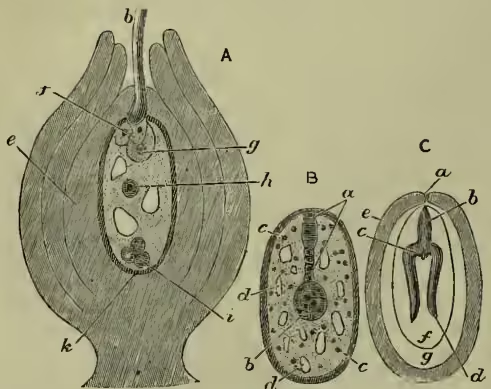
The embryo is the essential part of the seed; the other structures are subsidiary to its nurture, protection, and germination. Of these other parts of the seed the stored food material is of great importance, for after separation from the parent the embryo grows and sends out its rootlet, and pushes up its stem and expands its delicate leaves, in great part on the strength of what nutritive material it or the seed contains. This nutritive material, or 'albumen' as it is often called, out of which the first new parts of the young plant will be in great part built up, is formed after fertilisation—the physio- logical conditions are not understood—and is disposed within the seed in three different ways in relation to the embryo. It may lie in the cells of the nucellus, around but not within the embryo-sac, as in the seeds of bananas and ginger; then we call it perisperm. Or it may lie in more intimate relation to the embryo, within the embryo-sac (where a median nucleus seems to bear some relation to its formation), as in the seeds of wheat and castor-oil; then we call it endosperm. Sometimes there may be both endosperm and perisperm, as in water-lilies. But in many cases there is neither endosperm nor perisperm, all the food material being stored within the embryo itself—in its cotyledons—as in the pea and wallflower, apple and almond. Such seeds are somewhat confusedly called ex-albuminous.
The food materials, which are so often and by no means accurately summed up in the term albumen, vary in different kinds of seeds, but in a general way we may say that they consist of starchy, fatty, and proteid substances in varying proportions. In the development of animals they have their analogue in the yolk, and the analogy is especially true of the endosperm which is formed within the embryo-sac. But this analogy cannot be pressed, and it is perhaps more important to notice that the three ways in which the nutriment occurs—in the nucellus (perisperm), in the embryo-sac (endosperm), and in the embryo itself—form an intelligible series. The perisperm, which is least common, occurs in the nucellus—an outgrowth of the carpel—i.e. in the sporangium of the sporophyte generation (see FLOWER, FERN, OVULE). The endosperm occurs in the embryo-sac, has its centre in a central nucleus (formed from the fusion of two of the eight nuclei into which the primary spore-cell divides), and is therefore included in the prothallium of the gamophyte generation. Lastly, and most frequently, the nutrition may be stored within the embryo which results from the fertilisation of the egg-cell of the gamophyte.
Structure of the Seed considered in relation to the Protection of the Embryo.—As the embryo plant is a delicate structure, and as its separation from the parent plant is attended with vicissitudes, for the seed may be blown or carried into unfavourable conditions, and as most of the seeds which are liberated are not at once ready to germinate, it is important that the life of the embryo should be protected. This necessary protection is supplied in various ways. Around the ovule a double investment usually grows, and part of this investment is modified as the husk of the seed. The modification becomes marked as the seed grows ripe, as it begins to be ready for separation from the parent, as its supply of water begins to fail, as its vitality becomes more dormant; in fact the modification, which is a most useful one, is in part a necessary result of the physiological conditions of seed-ripening. In the modified husk it is generally possible to distinguish a thin, relatively unimportant internal tegmen from an outer, thicker, resistant testa. This testa may be leathery or woody, fleshy or gelatinous, smooth or hairy; it is variously adapted for the protection of the enclosed embryo. We may compare it to the shell or case which often surrounds the animal embryo. There is usually some relation between the nature of the seed-coat and that of the pericarp which surrounds the whole fruit; for in indehiscent fruits with tough walls (as in achenes and nuts) the seed-coat remains thin, while in dehiscent fruits the seed-coat is usually thick and hard. See GEOGRAPHICAL DISTRIBUTION.
Apart from the husk, the ripe seed is sometimes marked by a growth from its base or stalk. This is called the aril, and may be hairy as in willows and poplars, fleshy as in Euonymus, peculiarly shredded as in the mace of Myristica aromatica or nutmeg. When the growth from the base or from the funiculus of the seed is all on one side, it is called a caruncle, as in Chelidonium majus, Viola tricolor, and Ricinus communis. The outer surface of a liberated seed may also bear a mark at the place (hilum) where it separated from the funiculus; this is well seen in the common bean. Sometimes also a small opening or depression (cicatricula) persists as a remnant of the micropyle or aperture by which the pollen-tube passed between the coats of the ovule. In many cases the region marked by an external scar, corresponding to hilum or to micropyle or to both, is of special importance, the tissue of the seed-coat being modified so as to act as a sponge through which water soaks in to the embryo within. Many of the hard coats of seeds have, instead of this special sponge, numerous minute canals perforating their dense substance.
Structure of the Seed considered in relation to Seed-scattering.—Already in the article FRUIT reference has been made to adaptations which secure the dispersion of the seeds. Oftenest the dispersion depends upon the fruit, which may burst violently so that the seeds are scattered, or may be juicy and palatable so that it is eaten by birds and other animals, the indigestible seeds being in this way carried far and wide. Sometimes the fruit and the seed are virtually though not technically the same, as is the case in thistle-down, in which each little winged fruit borne about by the wind contains a single seed. Sometimes, however, the seeds themselves are borne about by the wind, as in willows and poplars, willow-herbs (Epilobium), cinchona, Aselepias syriaca, &c. The hairs which serve as parachutes to the seeds of poplars, willows, and the like are accessory growths outside the seed; a similar rich growth of cottony hair is characteristic of Gossypium and Eriodendron. It should also be noted that in many cases the seed is adapted to anchor itself, for the surface is often ridged or peaked, as in Hyoscyamus, Papaver, Nigella, or becomes glutinous in moist places, as in flax and some species of Plantago.
For further details as to the structure of seeds, consult morphological works such as Sachs's Text-book of Botany; for further details as to seed-scattering and the like, see
Kerner's Pflanzenleben, vol. ii. (Leip. 1891); for hints as to the practical study of seeds, see Strasburger's Practical Botany, trans. by Hilhouse (2d ed. 1889).
Vitality of Seeds.—Although the seed is a complex structure, it is usually able to remain for weeks, months, or even years in a state of dormant life hardly distinguishable from death except in its power of reawakening. We usually say that a seed is ripe at its separation from the parent plant, but this ripeness is not coincident with readiness to germinate. Sometimes indeed, as in the sheaves of corn in a wet season, the seeds may germinate before they leave the parent plant; those of ephemeral plants usually germinate as soon as they are scattered. In most cases, however, seeds are not ready to germinate until after they have remained for some time dormant. The seed of the mistletoe is ripe in autumn, but it does not germinate until April or May of the following year; that of Draba verna requires ten or eleven months of quiescence; that of Euphorbia cyparissias remains under ground from four to seven years; that of Euphorbia exigua is said to lie dormant for nine years. In some cases the reason of the prolonged postponement of germination is to be found in the hardness and thickness of the husks which surround the seeds, and in the slowness with which water seems able to penetrate into the dense cells; and it is likely that time is also required for ferments which the seeds contain to do their work of preparing the reserve food material for use in germination.
Seeds vary greatly in their powers of retaining their life. Those of the willows and poplars are peculiarly short-lived; unless they land in moist places they usually die in a few days. 'Seeds rich in ferment quickly lose their power of germinating. Ripe acorns will not germinate after a year, nor coffee-beans after six months. Nor do those which are rich in oil survive nearly so long as those whose reserve-products consist mainly of starch.' There is no doubt that carelessly-made experiments have given rise to a much exaggerated estimate of the powers of retaining vitality which seeds possess. Thus, we often hear of mummy wheat which has germinated after many centuries; but this assertion has not been substantiated. Indeed, modern experiments with the seeds of cereals show that they lose their vitality after ten years at most, and usually much sooner. Those of leguminous plants may, however, survive for several decennia. The vitality of seeds is also illustrated by the fact that some dry seeds can survive very high and very low (100° C. and -120° C.) temperatures. As to the real state of the protoplasm inside those seeds which remain so long without either living or dying, we are not yet in a position to make definite statements (see J. Wiesner's Biologie der Pflanzen, 1889).
Uses of Seeds.—As seeds are often rich in starchy, fatty, and proteid substances, their importance as food is very great. Those of cereals and leguminous plants are especially valuable; and the seeds of hazel, chestnut, cocoa-nut, Chenopodium Quinoa, Bertholletia, Pinus cembra, and many others are also edible. Others, such as mustard and nutmeg, furnish spices; many others, such as linseed, almonds, cocoa-nut, castor-oil plant, yield oil; cotton-wool surrounds the seeds of Gossypium, &c.; vegetable ivory is the dense endosperm of Phytelephas. Some seeds are rich in stimulants, as in coffee and kola plants; many, such as Strychnos nux vomica and Physostigma, yield drugs.