Ferns (Filices), as the most abundant and beautiful, most varied and typical class of higher (i.e. Vascular) Cryptogams, are of special interest alike to the botanist, the horticulturist, and the lover of nature. While Linnaeus included under his Filices all the Vascular Cryptogams, since Jussieu the equisetums, rhizocarps, club-mosses, and selaginellas have been separated off as distinct classes (see HORSETAIL, RHIZOCARPS, LYCOPodium, SELAGINELLA, ISOETES), as have more recently also the Ophioglossæ (see MOONWORT).
Since what we familiarly call the fern-plant is
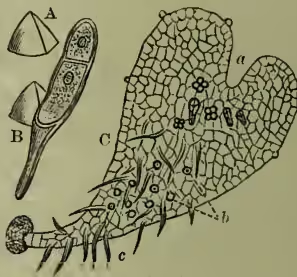
Germination of Prothallium :
A, spore; B, germinating spore; C, under surface of prothallium, showing archegonia, a; antheridia, b; rhizoids, c.
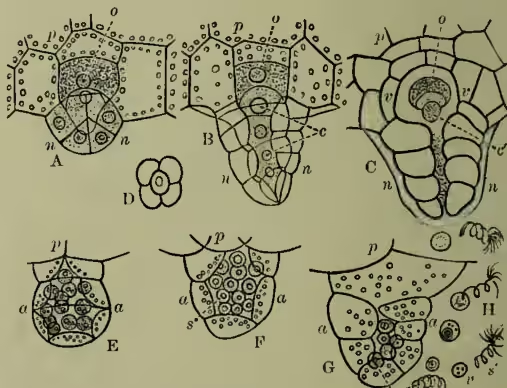
A-D, archegonia; A-C, longitudinal sections; D, transverse section of neck; p, prothallium; n, neck of archegonium; c, neck canal cells; v, venter of archegonium; c', ventral canal cell; o, oosphere. E-G, antheridia in transverse section; p, prothallium; a, antheridium; s, spermatocytes. H, spermatozoids, s', escaping from their vesicle, v, which contain starch grain.
only the spore-bearing member of an alternation of generations, it is convenient for the compre- hension of the life-history of the group, and still more for comparison with that of the other above-named Vascular Cryptogams, and of these with Gymnosperms (q.v.) and Phanerogams (q.v.), to begin with the familiar fern-spore, and trace its history round to the spore again, although necessarily only in the briefest outline. In fig. 1, A, this double-walled and characteristically formed and marked spore is represented; in B it is germinating into a short filament which soon broadens into a thin, flat, bilobed cellular expansion, the prothallus, C. This develops unicellular root-hairs from the under surface, and grows to a size varying on the average from th to th of an inch in diameter, resembling most nearly a small thallus of Liverwort (q.v.). It leads a perfectly independent existence, usually for several months, but sooner or later develops organs of sex, of which both are most commonly present upon the same prothallus. Near
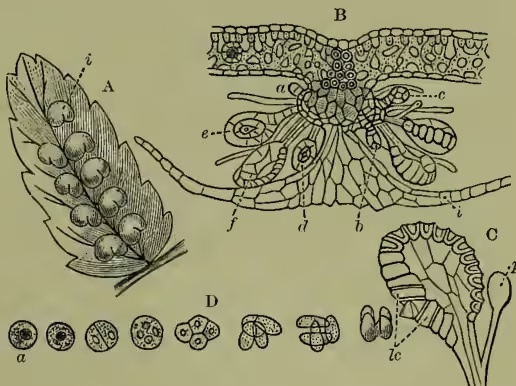
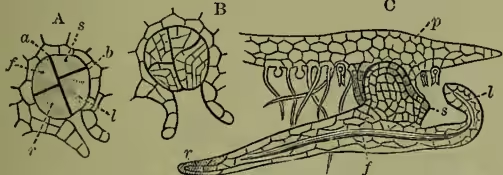
The male organs (antheridia) arise as epidermic papillae among the root-hairs, and segment into spheroidal groups of cells inclosing a large central cell, which soon segments into a group of smaller cells (see fig. 2, E-G). These undergo rejuvenescence, and escape in the ciliated state as free-swimming antherozoids, which (the prothallus growing in damp situations) find their way towards the archegonia. One antherozoid suffices to fertilise an archegonium, and, however many archegonia may be fertilised, the small prothallus is only able to bear a single fern-plant. The fertilised ovum segments into four portions (fig. 3, A-B) of which two segment to form the so-called foot, by which the young fern remains for a time attached to and supported by the parent prothallus (fig. 3, C); another goes on dividing to form the first root, while the remaining one gives rise to the stem and leaf. As the fern grows up it soon exhausts the prothallus, which dies off and disappears, and the fern gradually assumes the familiar vegetative development characteristic of the group.
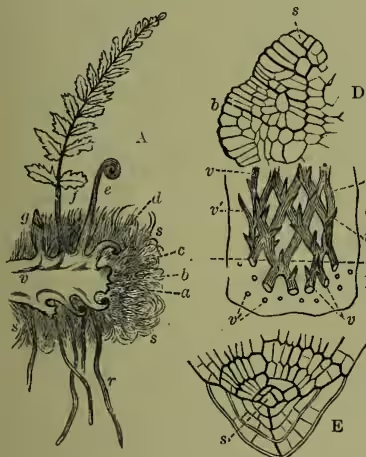
The fern-stem grows forward by an apical cell (fig. 4, D); the root-tip, too, has its apical cell, which also develops a root-cap (fig. 4, E). In the mass of embryonic tissue produced by the segmentation of the apical cell we soon distinguish (a) an external dermatogen layer which gives rise to the epidermis and its outgrowths, (b) internal strands of tissue of which many cells undergo characteristic modifications and give rise to the future fibro-vascular bundles, the whole lying imbedded in (c) the less modified ground tissue, which for the most part remains parenchymatous, but may in part become lengthened and hardened (as sclerenchymatous and prosenchymatous tissue), mainly for the purpose of increased mechanical strength. The fibro-vascular bundles form a meshwork anastomosing where bundles are given off into the leaves; they are closed and definite—i.e. contain no cambium and are consequently incapable of continued growth, save by addition in length at the growing-point. Their structure is concentric, the bast surrounding the wood. The axis forms most commonly a short root-stock; this, however, in some genera grows erect to a considerable height, giving us the characteristic and beautiful somewhat palm-like type of vegetation of the tree-ferns (see TREE-FERN). The leaves are developed in a close spiral succession just behind the growing-point, and are of slow growth, usually requiring two years to develop. Even in the expanded leaf growth may continue for a long time, the indefinite growth of the twining leaf of Lygodium being only the extreme case of this. Their characteristic circinate vernation—i.e. inrolled spiral or crozier-like folding in the bud—is very characteristic, and, as this must arise by a greater growth of the dorsal than the ventral surface, so the expansion of the leaf can be readily observed to be due to the temporary reversal of the disproportion, equilibrium being only reached with maturity. The leaf is sometimes entire, but is more frequently pinnate, bipinnate, or decompound: this arises, however, in a totally different way from the pinnation of the leaves of pteridophytes (see LEAF), being due to a dichotomy with unequal development of either fork alternately. The leaf-stalk or rachis is thus a false axis. The mode of origin is well revealed by reference to those tasselled varieties (fig. 6, A) which so frequently appear in widely distinct species in cultivation, exuberantly vegetative conditions admitting of the almost equal development of many successive secondary axes.
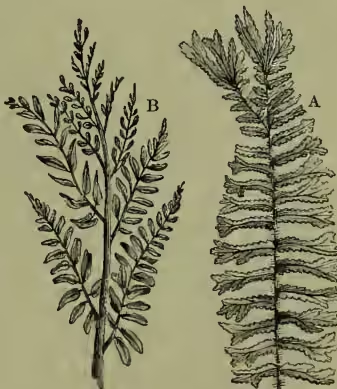
The process of asexual reproduction is in most ferns carried on by all the leaves indifferently (whence the attempt formerly so prevalent, but botanically unnecessary, to distinguish them from ordinary leaves as fronds). In many cases, however, this becomes restricted to particular portions of the leaf, apical in the common royal fern (Osmunda regalis, fig. 6, B), or median as in O. interrupta, or even to special fronds, as in the common hard fern (Blechnum spicant) or the parsley fern (Allosorus crispus). In such cases the vegetative development of the spore-bearing fronds is of course greatly checked, and we have a distinct foreshadowing of the stamens and carpels of the pteridophytes (although the spores have not as yet themselves acquired any character of distinct sex, much less the sporangia and the leaves which bear them). The fern may also exceptionally bud directly from the prothallus without archegonia (apogamy), or the prothallus from the frond without spores (apospory). Many ferns (particularly aspleniums) also multiply directly by budding upon the fronds.
At fig. 5, A, we have a fertile pinna of the common male fern (a name which of course refers merely to an old misinterpretation of the contrast of its rough foliage with that of the lady fern), showing the patches of spore-cases, or sori. The same figure (B) gives a section of a sorus, with developing sporangia under its epidermic cover or indusium. These are simply multicellular epidermic hairs, B, C, of which the central cell of the head segments and re-segments into spores (D). The spore-case is ruptured when ripe in various ways at once of practical interest and of high systematic importance, and the spores are thus diffused in great numbers to take their scanty individual chance of germinating as new prothallia.
Systematists vary somewhat as to the orders into which to divide this very large class, which includes about 70 genera and about 3000 living species, besides a great number of extinct forms. A convenient enumeration of these, however, may be taken as follows:
A. Sporangia thick-walled and without ring, arising in the leaf-tissue, and not as epidermal hairs (trichomes). I. Marattiaceæ—4 genera, with about 30 species in tropical America, Asia, and Oceania: Marattia, 10; * Angiopteris, 7.
B. Sporangia epidermal, with ring. (a) Ring indistinct. II. Osmundaceæ—2 genera, 11 species, scattered through all regions: Osmunda, 7 (see ROYAL FERN); Todea, 4. (b) Ring well developed; 6 orders, distinguished by details of sporangial structure—viz. III. Schizæaceæ—4 genera, tropical: Schizæa, 14; Antimia, 30; Lygodium, 30; Mohria, 2. IV. Gleicheniaceæ—3 genera, tropical, 40 species: Gleichenia, 8; Mertensia, 30. V. Hymenophyllaceæ (see FILMY FERNS)—3 genera, 200 species, chiefly tropical: Hymenophyllum, 80; Trichomanes, 90. VI. Cyatheaceæ, chiefly Tree-ferns (q.v.)—6 genera, with about 170 species, chiefly of southern tropical and warm temperate zones: Alsophila, 60; Hemitelia, 30; Cyathea, 40; Cibotium, 6; Dicksonia, 20. VII. Parkeriaceæ—2 genera, with only 2 species, Ceratopteris thalictroides, eaten as a vegetable in tropical Asia, and Parkeria pteridioides, found swimming in tropical American waters. VIII. Polypodiaceæ. While the preceding families contain only the genera above named, or at most one or two others, the Polypodiaceæ constitute the great majority (nine-tenths) of the whole group, and are widely distributed through all regions of the globe, although chiefly in warm climates. Of their families and leading genera a brief summary is therefore convenient.
(1) Sorus not covered by indusium. (a) Acrostichaceæ, with sporangia uniformly scattered upon the back of the frond or part of it—3 genera, 200 species, mostly tropical American, East Indian, and Australasian: Acrostichum, 170; Platycerium, 5, of which some species are cultivated, the stag's-horn ferns familiar in greenhouses. (b) Polypodiaceæ, or Polypodes (800 species), with sori variously arranged. (i.) Sori in streaks following the veins: Gymnogramme, 74, of which G. chrysophylla and G. tartarea, both West Indian, are cultivated as the Golden Fern and Silver Fern, names which well describe the characteristic beauty of the mealy wax which is so copiously secreted by glandular epidermic cells as to cover the lower surface. Sori in round or elliptical patches: Polypodium, 150, mostly tropical (see POLYPODY); Phegopteris, 70 (Beech Fern). Sori linear: Ceterach, of which the common Scaly Fern, C. officinarum, was long in repute as an astringent medicine. (ii.) Sori along the edge of the frond: may be continuous and uncovered (Notochlaena), or covered by a false indusium produced by the infolded edge of the frond, which is continuous in Pteris, 120 (see BRACKEN), discontinuous in Allosorus, 34 (Parsley Fern), while in Adiantum, 108 (see MAIDENHAIR FERN), the patches of sori are developed upon an inturned edge.
(2) Sorus covered by indusium. (c) Asplenaceæ, or Spleenworts (400 species), mostly tropical, with long or linear sori, with indusium arising laterally from above a vein: Blechnum, 50 (the Hard Fern); Asplenium, 300 (the Spleenworts); Scelopendrinum, 8 (see HART'S-TONGUE). Diplazium esculentum furnishes an edible rhizome in the East Indies. (d) Aspidiaceæ, or Shield Ferns (250 species), with dorsal sori of rounded shape, with shield-shaped or kidney-shaped indusia: Aspidium (Nephrodium), a large genus (220 species), including the Male Fern (see FERN, MALE) and many other well-known forms. Minor genera are Cystopteris, 9 (the Bladder Fern); Woodsia, 11; Struthiopteris, 3. (e) Davalliaceæ, with sporangia on a tooth, or in a furrow of the leaf edge—4 genera, with 200 species, almost all tropical: Davallia, 112.
* The numbers following generic names indicate the approximate number of known species.
Fossil Ferns.—We have a few fossil ferns even from the Silurian strata (Eopteris), while the Carboniferous rocks have yielded about 200 species, of which some were tree-like (Protopteris, Caulopteris, Pecopteris, perhaps also Neuropteris), while others (Sphenopteris, Hymenophyllites, &c.) appear to have formed great part of the undergrowth of the Carboniferous forest. Their remains also appear through the Secondary and Tertiary formations, approximating more nearly to those of our own day, and in the Tertiary indeed sometimes referable to existing genera (Alsophila, Lygodium, Osmunda). See PALEONTOLOGY (VEGETABLE). From the imperfect preservation of the sori and sporangia, the precise systematic position of fossil ferns can rarely with much certainty be determined.
Uses of Ferns.—Like other perennial stems and root-stocks, those of ferns contain a store of starch, and this becomes in many species of economic importance to communities which have not reached the agricultural stage. Foremost among these is rhizome of Pteris esculenta, the Tara (q.v.) of the Maoris, to which may be added Aspidium edule of Nepal, while the Cyathea medullaris in New Zealand, Angiopteris erecta in the South Seas, and other tree-ferns yield a kind of sago. Survivals of this use of ferns, or reversion to it in time of famine, also occur among more advanced peoples (see BRACKEN), and developing fern fronds are occasionally still eaten as a kind of salad in northern countries. Before the epoch of chemical manufactures the common ferns were a convenient source of potash, and their fronds are still often gathered as bedding for cattle. Their astringency gave them also a place in medicine, but the syrup known as Capillaire (see MAIDENHAIR) and the occasional use of male fern as a vermifuge alone survive in modern pharmacy. The characteristic beauty of their foliage has, however, given them in recent times a place in horticulture so important, especially in England, as to be sometimes compared to the historic Tulipomania (see TULIP) itself. The hardy ferns succeed well on shady rockwork, and the exotics under glass; their propagation, either by division of the root-stock (Davallia, &c.) or by sowing tolerably fresh ripe spores in saucers, kept moist and shaded under a glass (Pteris, Adiantum, &c.), is also easy. Many can be grown under a large bell-glass in rooms, or better in Wardian cases.
Various superstitions attach to ferns—the root of some kinds, for example, was in some places credited with the power of opening locks. By 'watching the fern' in solitude during the night of St John's Eve (24th June) one might hope to meet fairy benefactors, and receive a purse of gold; but the most widespread superstition is that fern seed, properly gathered, renders the bearer invisible. On the same eve of St John the fern puts forth at dusk a small blue flower, which soon disappears; and the wonderful seed, quickly ripening, falls at midnight, and should be caught in a white napkin. Fern seed is in some places a 'wish-seed,' enabling the bearer to attain his utmost wishes.
For guidance to the abundant literature on ferns, see Goebel's, Van Tieghem's, or other text-book of botany; the several works on ferns by Sir W. J. Hooker (British Ferns, Garden Ferns, Filices Exotice, Species Filicum); Heath's works (The Fern World, Fern Paradise, Fern Portfolio); also, for systematic studies, Smith's Historia Filicum; or general, for beginners, Moore's British Ferns. For information as to particular genera and species and their culture, see Nicholson's Dictionary of Gardening. See also De Bary (transl. by Bower and Scott, 1885).