Fishes, a great class of backboned animals, rising above the lowly vertebrates—Ascidians, lancelet, and round mouths—on the one hand, and leading on to the Amphibians on the other. The Tunicates or Ascidians (q.v.) are indeed numerous, but they are mostly degenerate; the level attained by the lancelet (Amphioxus, q.v.) is represented only by a single genus; the round mouths or Cyclostomata (q.v.) are few in number and partially retrogressive, but the fishes have been and are successful. In the possession of the characteristic vertebrate axis and central nervous system, in general compactness and co-ordination of structure, in their external armature of scales, and in their great fecundity, fishes have an easy pre-eminence over their invertebrate inferiors, while as successfully adapted forms they hold as firm possession of their own special medium as birds do of theirs. Their success may be read in the immense number of individuals, species, and genera, not only now, but in the great æons of the past; in the geological record which shows how the Cartilaginous Fishes (q.v.) or shark tribe have persisted strongly from Silurian times, or how the decadent Ganoid (e.g. sturgeon) order is followed by a yet richer predominance of the modern Bony Fishes (q.v.); and, furthermore, in the wealth of adaptive specialisation which distinguishes so many, and which in the case of the 'double-breathers' or Dipnoi prophesies, in the evolution of lungs, the important transition from water to terra firma.
Literary History.—The history of the study of fishes finds its beginning as usual in the work of Aristotle (384–322 B.C.), who distinguished over a hundred Ægean species, is perfectly clear in discriminating them from cetaceans and invertebrates, and records a surprising amount of information in regard to both their structure and habits. As with other departments of natural history, so ichthyology, in spite of its stable foundation, remained virtually unprogressive for eighteen centuries. In the 16th century at length Belon began to collect both specimens and information; Salviani, the physician of three popes, published an illustrated work characterised rather by artistic merit and popular interest than by scientific value; while Rondelet, as a medical anatomist, ventured further into structural details than either of his contemporaries. In the next century the classifying genius of Ray and Willughby achieved great progress, and the establishment of order was continued by Artedi, whose results were edited by his fellow-student Linnaeus. The labours of the Linnaean school were ably continued by Bloch and Lacépède, both working during the critical period of the French Revolution. The anatomical investigations of Alexander Monro, embodied in a classical folio, led on to the great Histoire Naturelle des Poissons begun by Cuvier, continued by his pupil and collaborateur Valenciennes, and finally left unfinished in 1845 at the twenty-second volume. The virtual discovery of the Ganoids by Agassiz, their investigation by Johannes Müller, the separation of the Dipnoi, the Cyclostomata, and Amphioxus by the same anatomist, and the discovery of Ceratodus, described in 1871 by Günther, were ichthyological events of great importance. Within late years, apart from the vigorous prosecution of anatomical investigation, the study of the development of fishes has yielded to such workers as Balfour, Parker, and Semper results of fundamental import; our knowledge of fossil forms is being continually enriched by the researches of Traquair and others; the theoretical aspects of structure have been especially investigated by Dohrn; while the high-water mark of general ichthyology is represented in the British Museum Catalogue of Fishes and other works by Günther.
General Survey.—After excluding the lancelet and the round mouths as survivors of far-removed ancestral forms, there remain four great orders of fishes properly so called—the Cartilaginous Fishes, the Ganoids, the Dipnoi, and the Bony Fishes or Teleosteans. The first three orders, of more ancient origin, are distinguished as Palæichthyes from the modern fishes or Teleostei which now form the vast majority. The cartilaginous fishes include the Elasmobranchs—skates and rays, sharks and dog-fish, with the addition of an unimportant sub-order, Holocephali, for the king of the herrings (Chimaera) and the genus Callorhynchus. The Ganoids or heavily-armoured fishes had their golden age in Devonian and Carboniferous times, and are represented nowadays only by a few genera—e.g. sturgeon (Acipenser), bony pike (Lepidosteus), Polypterus, and Amia; while the Dipnoi or double-breathers have only three representative genera, widely separated in geographical distribution—Ceratodus from Queensland, Protopterus from West and tropical Africa, and Lepidosiren from the Amazon. The Teleostean fishes include all the most familiar fresh-water and marine forms.
General Characters.—Like other vertebrates, fishes possess a segmented body, well indicated by the disposition of the muscles; a dorsal nerve cord, swollen anteriorly into a brain; a dorsal axis or notochord, more or less completely replaced by a genuine backbone; a set of respiratory clefts on the pharynx, which here persist and are associated with gills; a ventral heart; and eyes which arise for the most part as outgrowths of the brain. But their prominent peculiarities are associated with the three sets of structures—scales, fins, and gills. The scales are products of both outer and inner skin—i.e. of both epidermis and dermis; the unpaired median fins are always supported by skeletal rays, and the paired lateral fins or limbs are never in any definite sense fingered; the gills are retained throughout life, and except in the Dipnoi there are only the beginnings of pulmonary respiration. Technical negative characters are the absence of an allantois bladder and of a true inferior vena cava.
Structure and Functions.—The most characteristic form is that of herring, haddock, salmon, and the like, an elongated, laterally-compressed spindle, thinning off like a wedge posteriorly, and obviously the result of, or an adaptation to, the mode of aquatic progression. But the skates are flattened from above downwards, and the bony flat-fish, such as plaice, are compressed laterally; the eels, the ribbon-fish (e.g. Regaleus), the band-fish (Cepola), the pipe-fish (Syngnathus), are much elongated; while the globe-fish (Tetrodon, Diodon), coffer-fish (Ostracion), and sun-fish (Orthagoriscus) are more or less globular. The flat-fish live at the bottom; the elongated forms have a serpentine habit of creeping through crevices; the globular forms are sluggish, and may float apparently passive in the water. Prominent external characters, to be presently noticed in order, are the nostrils and eyes, the openings of the gill-cavities, the various kinds of scales, the paired and the median fins, the lateral sensory lines, the openings of the gut and urinogenital organs, the symmetrical or asymmetrical shape of the tail.
The skin has no true glands; the under stratum or cutis is without muscular elements; both layers take part in forming the exoskeleton of scales. In some bony fishes, in the Dipnoi, and in the sturgeon, goblet-like mucous cells burst and produce a slimy surface, but the usual slime so noticeable in skates and many other fishes exudes from special sensory tubes. Pigment cells under nervous control are common in the skin—e.g. in plaice, which thereby more or less rapidly alter their colour to suit their surroundings. The colours of fishes are often resplendent, and usually depend on the presence of lipochrome or fatty pigments, greatly enhanced by structural peculiarities and by the presence of a silvery foreground of guanine in the epidermic cells (see PIGMENT). As in the 'gemmeous dragonet,' the males are at the breeding season often transfigured in brilliant 'wedding' robes. The exoskeleton varies very widely, and so inconstantly that Agassiz' attempt to base a classification on the nature of the scales has been universally abandoned. Cartilaginous fishes are characterised by large or small skin-teeth, 'dermal denticles' or 'placoid scales,' forming the spines of a thornback or the shagreen of a shark. They are based in bone, cored with dentine, tipped with enamel, and are similar in structure and development to the true teeth of the mouth. The bone and dentine are formed from the cutis or dermis, the enamel from the epidermis. Skin-teeth occur here and there in other fishes, but flat scales predominate. These lie in sacks of the under skin, sometimes fuse together into plates, are covered with enamel in the bony pike and some other Ganoids, but are best known as soft overlapping plates in the common Teleosteans, where they are called cycloid or ctenoid, according as the free margin is entire or toothed.
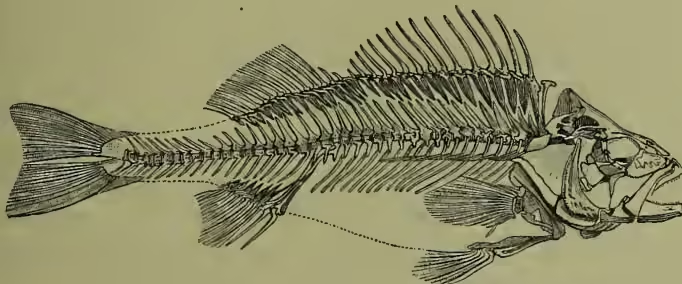
Skeleton.—The oldest vertebrate skeletons were cartilaginous, and the greater part of every individual framework has to recapitulate this stage. Very early, however, and at first in scales and teeth, bone was developed; but many of the older types of fishes are only slightly or very partially beyond the cartilaginous level. This is true of Elasmobranchs, Dipnoi, and some Ganoids, while other Ganoids and the bony fishes par excellence—the Teleosteans—have ossified skeletons. As in all other vertebrates, the longitudinal axis begins as a rod of cells on the dorsal wall of the gut. This notochord, itself of endodermic origin, becomes surrounded by a mesodermic sheath, and when a genuine vertebral column is established it is always by the predominance of the sheath over the enclosed rod. In some Elasmobranchs, in gristly Ganoids, such as the sturgeon, and in Dipnoi, the notochord with its sheath persists undivided; while in other fishes vertebral bodies, bony or otherwise, are established. The typical fish vertebra is concave at both ends, and every two adjacent vertebrae thus enclose a wide space in which the remains of the notochord persist. The bony pike (Lepidosteus), one of the bony Ganoids, stands alone in having vertebrae hollow behind but convex in front. The dorsal axis protects the spinal cord, which lies above, by forming a continuous tunnel, or by a succession of 'neural arches.' In the tail region 'haemal processes' similarly surround the blood-vessels, and the external ends of neural and haemal arches may come to be associated with dorsal and anal unpaired fins. The tail-end of the axis is usually bent upwards, but is straight in Dipnoi and a few others, and is surrounded by a special bony sheath in many Teleosteans. Thus, we have an asymmetrical tail in Elasmobranchs and some Ganoids, a perfectly symmetrical tail in Dipnoi, and a superficially symmetrical tail in most Teleosteans, and in some Ganoids, where the way in which the rays are disposed disguises the real upturning of the axis. When the notochord remains undivided the ribs are cartilaginous, and in all fishes they are very simple.
To understand the skull it is necessary to recognise its triple origin: (1) The brain is at first enclosed in a cartilaginous box, with which the gristly capsules surrounding nose and ear become at an early stage intimately associated. This chondrocranium or gristly brain-box is a truly axial portion of the skull, corresponding to a comparatively large number of segments (see SKULL). It is always well developed, and remains entirely unossified in Elasmobranchs alone, being in other fishes more or less modified into bones. (2) But, in the second place, what are called skin bones have to be taken into account. These dermal ossifications, 'membrane,' or better 'investing-bones,' probably had their historic origin in the fusion of bony scales. They are greatly developed in bony Ganoids and in Teleosteans, and more or less cover the chondrocranium on its roof, floor, and sides. (3) Lastly, there depend from the skull a series of arches, of which the posterior form supports for the gills, while the anterior form at least the groundwork of the jaws, and may enter into yet more intimate union with the main body of the skull.
Limbs.—The paired fins of fishes are in origin strictly comparable with the arms and legs of higher vertebrates, but their component parts are too simple to be very definitely compared with those of fingered or toed appendages, such as occur for the first time in amphibians. A typical fin consists of a basal piece, or of several, associated internally with a pectoral, and less constantly with a pelvic girdle. From the base a large number of rays radiate outwards. A most interesting type, from which some have attempted directly to derive the 'dactyls' of higher forms, is exhibited by the Dipnoi, in which a main axis runs down the limb (see CERATODUS). It seems most probable that the paired fins are concentrations or remnants of a continuous lateral fin extending along each side of the body, or of a series of limbs, one for each segment. Among the modifications worth noting are the enormous pectoral fins of skates, the anterior position of the pelvic fins in many bony fishes, and the entire absence of limbs in eels, pipe-fish, and a few other Teleosteans. As to their use, it must be remembered that the chief organ of locomotion is the tail, and that the paired fins serve to raise and depress the fish in the water, or to some extent as rudders. In a few cases, as in the climbing perch, they are utilised as scrambling appendages. The pectoral girdle which supports the fore-limb is cartilaginous in Elasmobranchs, Dipnoi, and some Ganoids, but is elsewhere ossified. Coracoid and scapular portions are distinguishable, while in Ganoids and Teleosteans there is a special development of investing bones. The pelvic girdle is a well-developed hoop in Elasmobranchs, while in Ganoids and Teleosteans, with one or two trifling exceptions, it is absent. The unpaired fins are derived from a continuous fringe seen in the embryo (see fig. 2), and this state occasionally persists. Usually, however, there are several dorsals and anals, and the great locomotor fin of the tail. The rays are horny, cartilaginous, or bony, and vary enormously in size, form, and number. Sometimes spine-like and all of one piece, sometimes soft and jointed, they afford valuable aids in classification.
Muscular System.—The great muscles of the trunk and tail, which do the work of locomotion, retain a segmented arrangement (as can be very plainly seen even on a boiled fish); and the segments or myotomes are separated by septa of fibrous connective tissue. Very remarkable modifications of muscular tissue form the various electric organs. See ELECTRIC FISHES.
Nervous System.—The brain and spinal cord are smaller in relation to the body than in other vertebrates. The growing skull outruns what it encloses, and the adult brain thus lies in a relatively large cranial cavity. All the usual divisions of the brain are developed, and in adult life lie virtually in one plane. Ten nerves from the brain, a large number from the spinal cord, and a sympathetic system exhibit their usual relations. See BRAIN, NERVE.
Sense Organs.—In most fishes a diffuse sensory system is located in the skin. Nerve endings of various kinds occur here and there over the surface, projecting superficially, arranged in definite lines, lodged in pits, or associated with a peculiar system of canals filled with mucus. The sensory structures thus variously arranged are distinguished as 'nerve eminences' and as 'end buds.' The former are very familiar in the lateral lines, one of which runs down each side of most bony fishes, and in the mucous canals which branch so abundantly on skates. The 'end buds' hardly occur in Elasmobranchs, but are abundant in Teleosteans, especially about the head. Near the mouth of the sturgeon, the cod, and other forms are tactile 'barbules.' Important too, though unfamiliar, are the 'eye-like,' 'pearl-like,' and 'phosphorescent' organs on the skin of deep-sea fishes.
The nose sacs lie ventrally in Elasmobranchs, in some of which—e.g. skates—they are connected with the corners of the mouth in a sort of hare-lip fashion. In Ganoids and Teleosteans the nasal sacs lie dorsally in front of the eyes, while the Dipnoi are alone among fishes in having posterior nasal apertures opening within the mouth, as in higher vertebrates.
The eyes (see EYE) are usually large, but are destitute of glands, and have but a slight development of lids. In fishes living in darkness, whether in caves, mud, or the deep sea, the eyes may be rudimentary (see BLIND FISH). The final position of both eyes on one side is a remarkable peculiarity of bony Flat-fishes (q.v.).
The ear, which is not very deeply buried, is surrounded by a cartilaginous capsule in Elasmobranchs and Dipnoi; but in other cases, whether the surroundings be gristly or bony, the wall next the skull is membranous. As in higher forms, the ear begins as an inturning of the skin, and a canal opening to the exterior in many Elasmobranchs persists as a remnant of this. From the comparatively superficial position of the ear, the waves of sound have not to penetrate so deeply as in higher vertebrates, and 'are conducted partly through the gill-cover (when present), and partly through the gill-slits or spiracle;' while in the skate there is what physiologically corresponds to a kind of drum. Inside the ear are the usual otolithic accumulations, sometimes soft, sometimes stone-like. In many Teleosteans, as may be conveniently seen in cod or haddock, one of the otoliths is strikingly large.
Intelligence and Emotions.—Fishes are not usually credited with many emotions or with much intelligence, but their natural life has not yet been sufficiently studied to admit of a fair verdict. As it is, anglers know how they become wary and cunning, and keepers of aquaria have recorded many instances of the commoner emotions and of fair intelligence. According to Romanes their emotions approach those of ants or of children four months old, except that no evidence of genuine sympathy is forthcoming. 'Fear and pugnacity are too apparent in fish to require special proof. The social or gregarious feelings are strongly shown by the numberless species which swim in shoals, the sexual feelings are proved by courtships, and the parental by those species which build nests and guard their young.' The stickleback woos his mate with undisguised passion and amusing coquetry, and guards the nest and eggs with jealous care. The Chinese butterfly-fish or Macropod, besides making an ingenious frothy nest of air and mucus, lifts the eggs into it by enveloping them in a cloud of bubbles. Tamed fishes have been known to become familiar with their keepers, and to answer to certain sounds. In natural life, the angling of the fishing-frog, the spitting of the archer-fish, the leaping of salmon and their careful selection of spawning grounds, the memory for localities which some are said to exhibit, all indicate that intelligence is not asleep in the fish's brain.
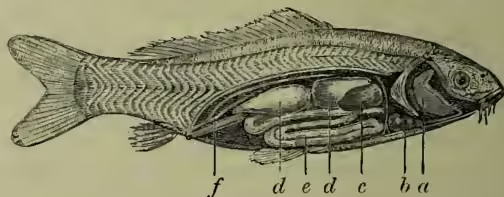
a, gills; b, heart; c, liver; d, swim-bladder; e, intestine;
f, ureters.
Alimentary System and Food.—The gut of fishes exhibits the usual vertebrate characteristics; but the distinctions of the several regions are often less marked than in higher forms. The mouth has no salivary glands, but, except in sturgeons, some Lophobranchs, and a few more, it has well-developed teeth on various bones. These mainly consist of ivory or dentine, tipped with enamel, and vary greatly in size, form, and mode of attachment. In cartilaginous fishes they rise out of a fibrous membrane which grows over the jaw as the outer teeth are worn away; in the angler (Lophius) and some other Teleosteans they bend backwards, and oppose the egress of struggling prey; in bony Ganoids and in Teleostei they occur on branchial arches (pharyngeal teeth), as well as on the usual bones of the skull. The mouth is ventral in Elasmobranchs, but in other cases is terminal, or occasionally upturned. The pharynx opens to the exterior by the respiratory gill-slits; gullet, stomach, intestine, liver, and gall-bladder have their usual relations; in Ganoids and most Teleosteans a large number of 'pyloric cæca' are given off at the beginning of the intestine; the pancreas is often diffuse or absent in Teleosteans; except in the last-named order, but especially in Elasmobranchs, there is a spiral ridge running down the large intestine, greatly increasing its internal surface. In Elasmobranchs and Dipnoi the gut ends in a cloaca or common chamber, into which the urinogenital ducts also open. In other fishes the anus lies in front of and separate from the other aperture or apertures.
The food of fishes is very diverse, from minute Protozoa to whales; they are carnivorous, vegetarian, or virtually omnivorous. Worms, crustaceans, insects, and molluscs and other fishes are important items in the carnivorous diet, in which they are usually voracious and often anything but discriminating. Some swallow mud for its living or decomposing contents; others, again, feed on sea and fresh-water weeds. The food is usually swallowed whole, though some sharks tear their prey into available pieces. Not a few forms manage to swallow other fishes out of all proportion to their own size, and in museums may be seen specimens which have managed to get outside victims even larger than themselves! Besides the formidable teeth of sharks and the like, the modifications of the jaws in sword- and saw-fish are conspicuous adaptations to a highly-evolved carnivorous habit.
Circulation.—In all fishes, except the Dipnoi, the heart receives impure blood only. This is driven by the muscular chamber or ventricle to the gills, whence being purified it is collected into the dorsal aorta which gives off arteries to all parts of the body except the head, which receives a direct supply from the most anterior of the vessels returning from the gills. From the body anterior and posterior veins convey the impure blood into the heart, into the so-called senns-venosus, or porch to the auricle. Though the direct work of the heart is solely to drive impure blood to the gills, it need hardly be pointed out that, as the chief origin of the alterations of pressure which determine the circulation, the heart has its usual, though obviously indirect, systemic significance. Except in the bony fishes, the ventricle is prolonged into a contractile tube known as the conus arteriosus; but, except in the Dipnoi, the heart is accurately described as two-chambered. Venous blood from the posterior region passes through the kidneys, forming a renal-portal system, as in amphibians and most reptiles, and there is the usual hepatic-portal system wherein vessels from the intestine pass into the liver. The hepatic veins leading from the liver enter the heart independently of the other posterior veins, and thus there is no strict inferior vena cava. In the Dipnoi the circulation has been modified with the acquisition of lungs. The heart receives pure as well as impure blood, and the auricle is divided by a septum. Fishes have the usual lymph system, and the ordinary blood-glands—spleen, thyroid, and thymus.
Respiratory System.—From the pharynx the characteristic vertebrate clefts open to the exterior, and admit of a water current in by the mouth, down the pharynx, out by the clefts. Between the clefts extend the supporting (branchial) arches, and these bear the folds or filaments on which the blood is exposed—in other words, the gills. In the cartilaginous fishes the separate gill-clefts are obvious enough externally; in other fishes the apertures are protected by a fold or operculum, and the partitions which form a series of chambers in skate or shark become more or less reduced. Thus, in the haddock there is one external opening on each side, and the gill-filaments are borne upon arches which are only attached at their extremities. The number of gills or clefts varies considerably; there are usually five in Elasmobranchs, four in Teleosteans. The primitive fish doubtless had a large number of gill-slits, and Heptanchus still survives with seven. In Elasmobranchs and some Ganoids the most anterior of the distinct gill-slits forms what is known as the 'spiracle,' opening on the top of the head, sometimes bearing a rudimentary gill, and lying in the position of the Eustachian tube in higher animals. In young Elasmobranchs, sturgeons, and many bony fishes there are to start with external gill-filaments, somewhat like those of a tadpole, while in Protopterus, one of the Dipnoi, there are three pairs of external gills in adult life. In the majority of fishes, but not in Elasmobranchs, nor in bony flat-fish such as plaice, there is an Air-bladder (q.v.) or swim-bladder, which arises almost always as a dorsal outgrowth from the gut, and propels the lung of higher vertebrates. As the name suggests, its function is in most cases hydrostatic, raising or lowering the fish according to its state of expansion or contraction, and it always receives a supply of arterial blood. In the 'double-breathing' Dipnoi, which gulp air at the surface, the swim-bladder functions as a genuine lung, and the same is true to a less marked extent of a few other fishes. In all Ganoids and in the Physostomi section of bony fishes the connection between air-bladder and gullet remains throughout life an open passage; in the other bony fishes (Physoelisti) the connection is a solid cord or is altogether obliterated. The sac extends along the back under the vertebral column, and is exceptionally (e.g. Polypterus) double like lungs; in four families of Teleostei it is connected anteriorly with the ear by a chain of bones, 'by means of which its relative fullness can be appreciated by the fish.' Some bony fishes—e.g. climbing perch and certain Siluroids—have curious accessory respiratory organs, and are able to remain for some time out of the water; and here also may be noted the dilatable sac which opens into the ventral side of the pharynx in some box or globe fishes—e.g. Diodon. See GILLS.
Excretory System.—To the zoologist the kidneys of fishes are of great interest, because it has been possible through the study of their development to reach an approximate elucidation of the 'homologies' or real resemblances in the different parts of the urinogenital system throughout the vertebrate series. Only two general sentences are here permissible. At an early stage a segmentally arranged series of ciliated excretory tubules, or nephridia, make their appearance; they are strictly comparable to the excretory tubes of invertebrates, and a certain unity is thus demonstrable between earthworm and dog-fish. Of these little tubes some form the adult kidney, others the 'Wolffian body' or 'parovarium,' while others disappear. But, besides the tubules which lie transversely, there is on each side a longitudinal duct with which some at least of the nephridia become connected; this so-called 'segmental duct' is either derived, as the insinking of a skin (ectodermic) groove, or else from the external portion of the middle (mesodermic) layer; on each side it typically becomes double, and forms the reproductive ducts for either sex, but the half which is important in the male is rudimentary or unimportant in the female, and vice versa.
Reproduction.—The sexes are almost always separate, but a male organ or testis 'is constantly found imbedded in the wall of the ovary in Chrysophrys and Serranus (two bony fishes), and the last-named fish is said to be self-impregnating.' As in most other vertebrates, occasional hermaphroditism occurs as an abnormality—e.g. in mackerel and herring. The males of bony fishes are usually smaller than their mates, and sometimes very markedly so; in many cases, especially at the breeding season, they are further distinguished from the females by bright colouring. Thus, the male 'gemmeous dragonet' (Callionymus lyra) is so flushed with brilliant colour that it was regarded by Linnaeus as quite distinct from the 'sordid,' dingy female. The male sea-scorpion (Cottus scorpius), usually ill-favoured, becomes gorgeously adorned during the spawning season, and the same is true of a great many forms. In Elasmobranchs the males are distinguished by the possession of copulatory organs or 'claspers.' As the males often fight with their rivals, special adaptations have come to be associated with this habit; of these the hooked lower jaw of the male salmon, so suitable for butting, is perhaps the most familiar instance. In many marine fishes the females appear to be in a very large majority. There are frequently other differences between the sexes, especially as regards the unpaired fins, and Darwin also quotes some cases (especially Ophidium) where it seems that the males, and the males only, have the power of producing sounds. See SEX, SEXUAL SELECTION. The ovaries and testes are almost always paired organs, which become large and conspicuous in the reproductive season. In all Teleosteans save one, and in the bony pike (Lepidosteus), there are ducts continuous with the ovaries; in other fishes the eggs burst into the body-cavity, and are subsequently pressed into the ducts. The male elements are likewise occasionally set free in the body-cavity (Dipnoi and Murenidae), but usually pass more or less directly into ducts. In most Elasmobranchs, in Ganoids, Dipnoi, and a few Teleosteans, there are two openings or 'abdominal pores' from the body-cavity to the exterior, which perhaps represent a primitive exit for the reproductive elements, and in a few cases still serve for that purpose. In the great majority, however, the reproductive ducts fulfil their usual functions, and open either into a cloaca in Elasmobranchs and Dipnoi, or behind the anus in Ganoids and Teleosteans.
Most fishes are oviparous, laying the eggs as such, and that usually in great numbers. They are extruded on the bottom—e.g. among gravel—or in many cases are discharged broadcast in the water. A few fishes, such as sturgeon and salmon, ascend rivers for spawning purposes; while others, like the eel, descend to the sea. The female salmon is careful in the choice of a spawning ground, and in ploughing a groove for the eggs among the gravel is said to avoid areas which have been already occupied by her forerunners. In regard to trout, it has been observed that the absence of a suitable bottom may lead to retention of the ova, to consequent disease in the organ, and to an inferior brood the next season. The eggs of oviparous Elasmobranchs are surrounded by a chitinous case, the so-called mermaid's purse, so often found empty on the beach. This sheath is a product of the lower portion of the oviduct, and often has tendril-like prolongations at the corners, which twine themselves automatically round seaweed, where the embryos are rocked till the time of hatching. Most sharks and a few Teleosteans bring forth their young alive. In the viviparous Teleosteans the eggs develop in the ovary; in the sharks, in the lower portion of the oviduct. Two of the viviparous sharks (Mustelus levis and Carcharias glaucus) are of peculiar interest in the establishment of a vascular placenta-like union between the yolk-sac and the wall of the oviduct, an arrangement which has also been noticed in a couple of lizards. In the Teleostean Anableps the yolk-sac has villi which protrude into a dilated ovarian chamber and absorb nutritive material. As to fertilisation, the usual process is that the male deposits the spermatozoa or the milt upon the laid eggs or spawn, but it is evident that fertilisation must be internal when the forms are viviparous or when the eggs are enveloped in a firm sheath.
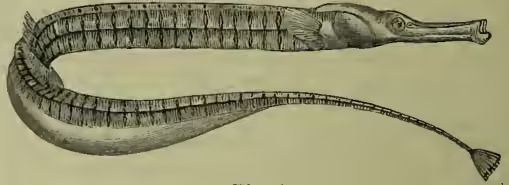
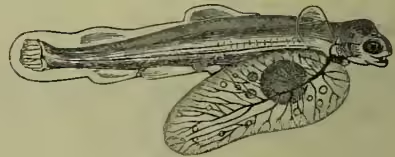
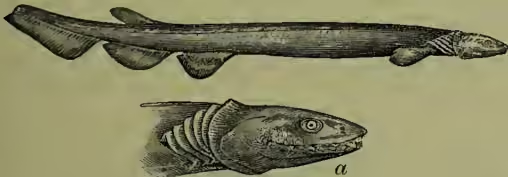
The male of a Pipe-fish (Syngnathus acus), with pouch beneath the tail for carrying the eggs (after Günther).
Parental Care is but rarely exhibited by fishes, and then usually on the part of the males. Love for offspring usually varies inversely with their number, and those of fishes are legion; nor would young fish benefit much by any care, except an impossible providence which would prevent their being swallowed. Apart from numerous viviparous forms, the female of a Siluroid genus (Aspredo) carries the ova on her belly till they are hatched, just as the Surinam toad bears her progeny on her back; and in Solenostoma a pouch for the eggs is formed by the ventral fins and skin. The male stickleback (Gastrosteus) makes a nest of grass stems and water weeds, weaves it round with mucous threads (which seem to be almost pathological products of the kidneys compressed by the enlarged male organs), and jealously guards the eggs laid by his mate. The same is said to be true of a species of Chromis from the Sea of Galilee; and more or less elaborate nests are formed by the males in the genera Cottus, Cyclopterus, Antennarius, Ophiocephalus, and Callichthys. 'The male of some species of Arius carries the ova about with him in his capacious pharynx,' while in the pipe-fishes and sea-horses various abdominal brood-pouches are developed.
Development.—The fish egg is a small thing, except in the cartilaginous fishes, where it is very large, and enclosed in the characteristic chitinous case. In other fishes there are only the usual egg-sheaths, such as the zona radiata, with one aperture or more for the entrance of the spermatozoon. Fertilisation usually takes place in the water, much less frequently within the female. Since the old experiments of Jacobi (1757-63), artificial mingling of the sperms and eggs has often been resorted to, both in the embryological laboratory and in the practical work of stocking rivers (see PISCICULTURE). In the egg the yolk usually bears a large proportion to the strictly formative material, and the division with which development begins is partial in all known cases except the sturgeon and the bony pike. Development is, in most cases, a rapid process, and the newly-hatched forms bear the still incompletely absorbed yolk-sac dependent from their ventral surface. The larval forms are often unlike the parents, for instance, in the possession of external gill-filaments in Elasmobranchs, &c., or in minor features connected with mouth, fins, and armature. The most striking change is that which happens to young, bony Flat-fish (q.v.). See EMBRYOLOGY, and such articles as SALMON.
Relation to Environment.—In many fishes the limit of growth is less definite and less rapidly attained than is usual among higher vertebrates. In favourable circumstances individuals will go on growing, and may far outstrip the average of the species, while, on the other hand, a confined habitat and restricted food-supply may reduce the size. 'The young of coast-fishes, when driven out to sea, where they find a much smaller supply of food, remain in an undeveloped condition, assuming a hydropic appearance.' Such, probably, are the enigmatical Leptocephalidæ. Besides affecting the size, the nature of the surroundings and the nutritive supply influence both form and colour. It is a well-known fact that the same species of trout presents a variable appearance, according to the habitat of the individual. It rarely happens among fishes that the pigment wholly disappears, but it is not uncommon for an 'incipient albinism' to be produced by a change of the contents of the pigment-cells into yellow, a variation most familiar in the gold-fish, but occurring in not a few. Very different, and of more importance in the life of the fish, is the power which some possess of sympathetic change of colour. Thus, in the place the colour of the environment influences the eye, a nervous impulse determining the contraction or expansion of the pigment-cells passes to the skin, and the animal is adapted to the colour of the ground on which it rests. Few general statements can be made as to the sensitiveness of fishes to changes in the medium. Pelagic fish are very susceptible; but other marine forms—e.g. flounders—thrive well in fresh water. 'A shark lives in Lake Nicaragua; a goby, blenny, and Atherina in the lakes of North Italy.' The passage of a fresh-water fish to the sea is very rare. Many fishes can endure prolonged fasting and also lowered temperature, but in both these respects marine forms are far excelled by those inhabiting fresh water. Low temperature causes some fishes to pass into a torpid state of 'Latent Life' (q.v.), or Hibernation (q.v.), while in the dry season of tropical countries other forms (e.g. Siluroids and Protopterus) encyst themselves in the mud, where they may remain like living mummies for prolonged periods. See also BLIND FISH.
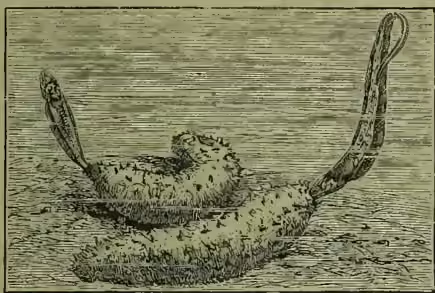
Some small fishes live in amicable partnership with other animals—e.g. inside large sea-anemones (see COMMENSALISM). The genus Fierasfer seems in a fair way to become parasitic, since the members find more or less permanent lodging within sea-cucumbers, medusæ, and other animals, feeding, however, not upon their hosts, but upon the minute animals which they contain. Fishes themselves are infested by numerous internal and external parasites. Crustaceans, or Fish-lice (q.v.), occur in great abundance on skin and gills, especially on sluggish or on sickly forms. Various flukes are also found externally, and a large number of Cestodes in tapeworm or bladderworm stages live in the viscera. The pike and burbot are hosts of the immature stages of the formidable Bothriocephalus; the stickleback often bears the young of the fresh-water mussel; a peculiar hydroid (Polypodium) is parasitic on the eggs of the sturgeon; and so on through a long series (see PARASITISM).
Distribution in Space.—Most fishes live either exclusively in the sea or exclusively in fresh water, but some pass for spawning purposes from the one to the other, and not a few thrive with apparent indifference in either or in a brackish medium. The marine forms enjoy a wider distribution over the globe, but the inhabitants of fresh water also spread by circuitous swimming, by changes in the river systems, and even by short overland marches, or by having their ova carried on the feet of water-birds. Both marine and fresh-water forms occur in Polar regions. Günther describes about forty species from the icy waters at or beyond the 1500 fathom line; and fishes are still present in Alpine lakes about the level of perpetual snow, up, indeed, to a level of 15,000 feet or more. Of genuine fresh-water fishes about 2300 species are recorded, of which four are Dipnoi, thirty-two Ganoids, and the rest Teleosteans, with a marked predominance of carps or Cyprinidæ, and Siluroids or cat-fishes. The marine forms are usually distinguished as shore, pelagic, and deep-sea fishes. The shore fishes, including about 3500 species, frequent the coasts, and rarely descend below 300 fathoms. The pelagic fishes swim and usually spawn in the open sea, are less numerous than the shore forms, and thrive especially in the equatorial waters: the cartilaginous fishes form a large percentage of the pelagic fauna. The deep-sea fishes include over a hundred genera, most of which are Teleosteans. Of these forms, monographed by Günther in his Challenger Report, a little more must be said.
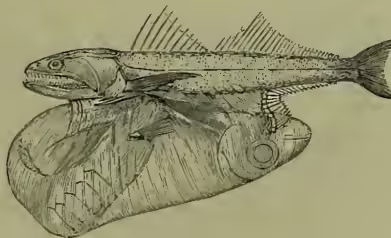
Deep-sea Fishes were not known with any definiteness till the Challenger expedition virtually discovered the new zoological world which they inhabit. The explorations of the Talisman and the Travailleur have also contributed important results to our knowledge of these usually strange-looking forms. To understand their peculiarities it is necessary to recall the principal factors in their environment. (a) Sunlight is absent, and so often are eyes, which, in the absence of stimulus, remain undeveloped. It is interesting to notice Günther's results. From a depth of from 80 to 200 fathoms the eyes are larger than usual, to make the most of the scanty sun-rays which fade away at those depths. Beyond the 200 fathom line 'small-eyed fishes as well as large-eyed occur, the former having their want of vision compensated for by tentacular organs of touch, whilst the latter have no such accessory organs, and evidently see only by the aid of phosphorescence. In the greatest depths blind fishes occur, with rudimentary eyes, and without special organs of touch.' The Phosphorescence (q.v.) is produced by numerous marine animals and by the fishes themselves. Many of the latter exhibit round, shining, mother-of-pearl-coloured bodies, imbedded in the skin, in part sensory, in part phosphorescent. (b) The temperature of the bottom is low and equable, and a wide range of distribution is thus unimpeded. (c) The pressure is, of course, enormous at those great depths, and, though the fishes feel it as little as we do that of the air, it has doubtless been an important factor in determining their structure. Bones and muscles are less developed; the former are very light and brittle, the latter are thin. The forcible ascent to diminished pressure, of course, produces rupture and other changes, so that it is sometimes difficult to infer the natural form of the body and texture of the tissues. (d) As sunlight is absent, so also are plants. The abyssal fish are therefore carnivorous, 'the most voracious feeding frequently on their own offspring, and the toothless kinds being nourished by the animalcules which live on the bottom, or which "like a constant rain" settle down from the upper strata.' Some of the carnivorous forms show evidence of great rapacity, and not a few are characterised by wide gape and dilatable stomach, able to 'receive a fish twice or thrice the bulk of the destroyer.' The skin in many deep-sea fishes is marked by an extraordinary development of the mucus-secreting system, and this exuded layer has been observed to be phosphorescent in some fresh specimens. The colouring is simple, most of the forms being either black or silvery. The absence of light and the sluggish habit may have something to do with this peculiarity. (e) Lastly, the perfect calm of the abyssal water must be remembered, and associated with the tendency many of the fishes have to develop long, delicate filaments on fins or tail. As to the maximum depth at which fishes certainly occur, Bathyophis ferox was dredged from 2750 fathoms.
Classification.—The chief technical characteristics of the four orders, Elasmobranchii, Ganoidei, Dipnoi, and Teleostei, may be summed up in the following table; but the articles BONY FISHES, CARTILAGINOUS FISHES, DIPNOI, and GANOIDS should also be consulted at this stage, as well as special articles on distinctive types.
| Elasmobranchs. | Ganoids. | Dipnoi. | Teleosteans. | |
|---|---|---|---|---|
| SKIN ..... | With skin-teeth. | Large hard scales. | Large soft scales. | Usually soft scales. |
| LIMBS ..... | Large; hind pair abdominal. | Hind-limbs abdominal. | With a median axis. | Hind-limbs often far forward. |
| TAIL ..... | Asymmetrical. | Asymmetrical, or the same disguised. | Symmetrical. | Apparently, not really, symmetrical. |
| SKELETON ..... | No investing bones; almost wholly cartilaginous. | Investing bones on skull and pectoral girdle; otherwise cartilaginous or bony. | Persistent undivided notochord; largely cartilaginous. | Well ossified, with completely formed vertebrae, and well-developed investing bones. |
| OPTIC NERVES.... | Fuse with a crossing of fibres near the brain. | The same. | The same. | Cross without fusing. |
| GILL-APERTURES... | 5-7 opening externally, usually + a spiracle. | A cover or operculum; sometimes a spiracle. | Covered internal gills, in one case + external. | An operculum. |
| SWIM-BLADDER .... | None. | Present with persistent duct. | Functional as a true single or double lung. | Present, with or without persistent duct. |
| INTESTINE ..... | Cloaca; spiral fold in intestine. | Anus separate from urino-genital aperture; a spiral fold. | As Elasmobranchs. | No spiral fold; separate anus, and often separate urinary and genital apertures. |
| HEART ..... | Contractile conus arteriosus. | The same. | The same and a septum between auricles. | No contractile conus. |
| OVA ..... | Few, large, internally fertilised eggs; many forms viviparous. | Small, externally fertilised; oviparous. | Numerous small eggs; oviparous, but little known. | Numerous small eggs, externally fertilised, except in a few viviparous forms. |
| ALIA ..... | Males with copulatory organs on hind-limb. | A decadent order with seven or so living genera. | Nostrils communicating with mouth. Three living genera transitional to amphibians. | The vast majority of living fishes. |
In addition to the four orders above contrasted, it is usual to distinguish a fifth—Holocephali—for the reception of Chimaera (q.v.) and Callorhinchus. These two forms, of which no notice has been taken in the above general sketch, agree in most of their features with the Elasmobranchs, but differ, for instance, in the development of an operculum, in the distinctness of the anus from the urinary and genital apertures, and in the nakedness of the skin.
History of Fishes.—The earliest fish remains occur in the Upper Silurian strata, mostly in the form of spines, scales, shields, and mere fragments.
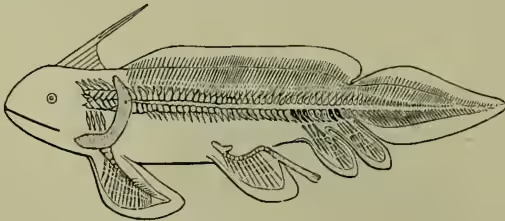
Some of these belonged to true sharks, but the armoured Ganoids (Pteraspis) were also alive in that ancient time. In the Devonian period both orders were represented, but the Ganoids greatly predominated, if the remains may be trusted as affording a true index. In the Carboniferous the Ganoids continued their long golden age, and the Elasmobranchs were also abundant. We figure a remarkable form (Pleuracanthus) recently described from the French coal-measures, which the discoverer, M. Brongniart, regards as representing at least a distinct order, perhaps a sub-class, the 'Pleuracanthides'—'a group ancestral to and connecting the Dog-fishes, Cestracions, Rays, Chimaeras, Sturgeons, and Ceratodus.' Very singular and novel are the two small anal fins, which are 'placed one behind the other, and have the appearance of true limbs.' In the Jurassic strata the modern types of shark and dog-fish become numerous, and have so remained ever since. Far otherwise the Ganoids; relatively few survived the Jurassic epoch, and the few genera now living form but a paltry remnant of a vast and heterogeneous host. Some of the ancient Ganoids exhibit characteristics which link them to the Dipnoi, and it is generally admitted that it was from Ganoidei ancestry that the Dipnoi (which appear in the Permian) had their origin. The occurrence of the modern genus Ceratodus in strata so far back as the Permian 'presents the most remarkable instance of persistence in the whole range of vertebrate animals.' The Dipnoi form a transitional order, leading on to amphibians, and probably never included more than a progressive few. In another direction, but probably also from Ganoidei ancestry, the modern bony fishes or Teleostei evolved. They appear for the first time in Creta- aceous strata, 'being immediately preceded by the Teleostoid group of Ganoid fishes of the family Leptolepidæ, which effects a passage to them' (Heilprin). Forms with open swim-bladders, and forms in which the connection with the gut is closed, both appear early, but there is no doubt that the former (Physostomi) are the more primitive. Herring, smelt, pike, and other bony fishes of to-day occurred in the chalk period, but it was only in the Eocene or early Tertiary times that the modern fauna became established. Fresh-water forms must be regarded as derived from marine forms, and the change of habitat probably began in very early times.
a, head on larger scale.
To sum up—the first fishes were probably somewhat after the shark pattern; these were soon followed and outnumbered by Ganoid forms; in successive æons the Elasmobranchs increased and remained firmly established if not markedly progressive; the Ganoids on the other hand waned most markedly, but according to many they gave rise to the highly successful Teleostean type, and to the progressive transitional Dipnoi. At present the Teleosteans are, of course, in an overwhelming majority; next come the persistent Elasmobranchs, including Chlamydoselachus, a Japanese shark which is said to be the oldest living type of fishes, with direct ancestors of Devonian antiquity; the eight or so living genera of Ganoids are a mere handful compared with the extinct forms; while the Dipnoi are only represented by three genera. As to the relations of fishes to the round mouths or Cyclostomata, to the lancelet or Amphioxus, to the Ascidians or Tunicata, and to still more remote worm-like forms such as Balanoglossus, it is only possible at present to say that all are offshoots from primitive vertebrate types.
Economic Interest.—Apart from the import of fishes in the general economy of nature, their practical interest for mankind is centred in the fact that so many form part of our food-supply. Their importance in this respect is especially great in Polar regions, where other sources of nutrition are scanty. It may be here noticed that many are temporarily or constantly poisonous. The poison is in the blood and is destroyed by heat in the eels; it is constant and in the flesh in Clupea venenosa, various globe-fish, and many others; it is temporary and in the roe of spawning pike, burbot, &c.; while in many cases it is not intrinsic, but due to the food on which the fish lives, or to decomposition products formed after death. Gills and other parts which quickly decompose should always be avoided. Some fishes, such as stingrays and weavers, readily give wounds with their spines, and the mucus of the body thus inoculated produces symptoms of poisoning. In Synanceia, Thalassophryne, and other forms there are true poison organs associated with spines, and the results of a wound are sometimes fatal. Besides the great nutritive utility of fishes, there are a few minor products of some importance, such as cod-liver oil from the liver of Gadoids, isinglass from the swim-bladder of sturgeons, &c., and shagreen from the skin of sharks and rays (see PISCICULTURE).
From their position near the base of the vertebrate series fishes are the most hopeful subjects for the solution of many of the fundamental problems of zoology; in their endless variety of habit they afford wide scope to the observant naturalist; while for their grace of movement, beauty or quaintness of form, and splendour of colouring they merit a larger share than they have hitherto received of artistic attention.
BIBLIOGRAPHY.—General: A. Monro, The Structure and Physiology of Fishes (Edin. 1785); Cuvier and Valenciennes, Histoire des Poissons (22 vols. Paris, 1828–49); Von Siebold and Stannius, Handbuch der Zootomie (Berlin, 1854); Hubrecht and Sagemehl, Brown's Thierreich (in progress); general works of Owen, Huxley, Gegenbaur, Wiedersheim, &c.; Günther, Introduction to the Study of Fishes (Edin. 1880); British Museum Catalogue of Fishes (1859–70); Challenger Reports, especially Deep-sea Fishes; Rolleston and Hatchett Jackson, Forms of Animal Life (Oxford, 1888). British fishes: Day (1884), Couch (1862), Yarrell (1841). American fishes: Bulletin of United States Fish Commission. Development: see literature at EMBRYOLOGY. Theory of structure: especially Dohrn, Mittl. Zool. Stat. Neapel. Fossil fishes: L. Agassiz, Poissons fossiles (1833–44); Zittel's Palæontologie (1887). For practical study: Parker's Zootomy (1884); Marshall and Hurst's Practical Zoology (2d ed. Lond. 1888).—The whale and the sturgeon, either when thrown on shore or when caught near the coast, are at common law the property of the crown, and as such are known as Royal Fishes.