Fruit. In popular language, the term fruit is very vaguely employed. When extended beyond the common limitation of usefulness to man or beast, it tends to be applied to any plant-structure, phanerogamie or cryptogamie, which contains the germ of the new individual—to all the organs of the germ of the new individual—in short. But, as common observation deepens into botany, we find ourselves gradually led to the more precise restriction of the term fruit to the ovary of angiosperms (monocotyledons or dicotyledons) after fertilisation (see FLOWER, OVARY).
The numerous and interesting adaptations of different fruits to the preservation and distribution of the seed will be more conveniently outlined under SEED, while the periodic rhythm between vegetative and reproductive growth to which the question of fruit attracts our attention must be discussed under the more general head of REPRODUCTION. The special structure and physiology of fruits here remain to be considered.
Since the dawn of modern botany, the multifarious forms of fruit have led to many attempts at their classification. Yet the student is more apt to be overwhelmed by the resulting disorderly and redundant nomenclature of the subject than impressed by its systematic clearness. If, however, we keep fast hold of the elementary conceptions of vegetable physiology, morphology, and evolution, the difficulty of enumerating and classifying the various forms of fruit becomes greatly diminished. We must of course assume a knowledge of the general morphology of the Flower (q.v.).
Starting then with those simplest flowers in which all the carpels are separate, we find the stigma and style usually withering back as no longer of service, and the ovary enlarging, as the fertilised ovules grow up into seeds. But in many such simple flowers more ovules are produced than are fertilised, and generally also more fertilised than can be developed up to maturity; hence the reduction of the ovules is exceedingly common. The alternative of reducing the number of carpels also commonly appears: hence in the same order of Ranunculaceæ we have on the one hand the anemone with its multitude of small ovaries which only mature a single ovule, and on the other the larkspur or monkshood with few carpels, but these many-seeded. This process of reduction of the number of carpels or ovules, or of both, has not only taken place in the process of past evolution of the great majority of plants, but is still frequently to be observed in the development of the individual, as is well seen by comparing the characteristically one-celled and one-seeded acorn with a section of the three-celled and six-ovuled ovary from which it actually arose in spring, or, more simply, by recalling to memory the abortive ovules and the corresponding abortion of one or two of the original three divisions of the ovary in the fruit of the horse-chestnut.
A second common-sense 'principle of fruit-making,' as we may call it, is reached through keeping clearly in mind the nature and origin of the ovary; for, however the upgrowth of the axis may in perigynous or epigynous flowers conceal this (see FLOWER), we know the ovary primarily to have arisen from one or more carpellary leaves, of which the individual development has been so greatly checked (doubtless through the precocious development of their sporangia—i.e. ovules), that so far from becoming expanded like all other appendages, they remain closed upon the ovules, and frequently even coalesce with each other from the base upwards, so forming a many-celled ovary, often even with united styles or even stigmas. Yet the tendency to their individual expansion is not lost; in many monstrosities, and normally a few types, such as the common mignonette, the carpellary leaves early begin to expand, so opening the ovary and exposing the seeds long before ripeness. Far more frequently, however, this final development of the carpellary leaves is delayed until the growth-processes of the seed and fruit have ended, and it is therefore accompanied, or even preceded, by their death; the separation often indicating the lines at once of leaf-margin and leaf-fall.
In the best developed carpellary leaves, such as those of the more floral Ranunculaceæ, we naturally find the ovary 'dehiscing along the ventral suture'—in more simple and less empirical language, the carpellary leaf opening along the line of its united ovule-bearing margins. This is what is termed a follicle (fig. 1, f).
Since, however, the ovules are on the united margins, the midrib tends to become mechanically unimportant, and to interpose little or no resistance to a tendency to split or tear along its fold, as well as to open along the united margins. Such 'dehiscence by both dorsal and ventral suture' gives us the modification of the follicle known as a legume or pod (fig. 1, e).
A very familiar type, which must not be confused with the pod, is the silqua (or when shortened and broadened the silicula) of Cruciferae. Here the placental edges of two united carpels develop a transverse septum which divides the fruit (fig. 1, d); and this is left when the lobes split away, as so familiarly in Honesty.
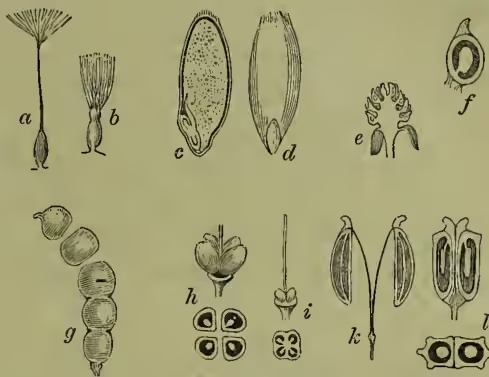
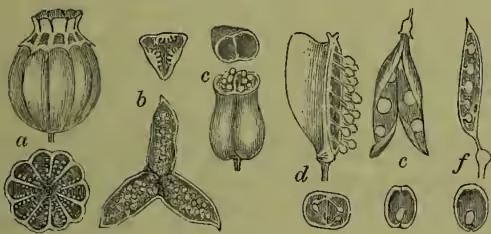
Among united ovaries which readily split open at the united margins (septicidal) we may note that of Gentian (q.v.), while the more familiar three-celled ovary of a violet (fig. 1, b) or rock rose with its parietal placentation gives a characteristic example of dehiscence along the midribs of the united carpels, so opening the loculi (loculicidal). In the five-celled capsule of the Geranium (q.v.) the carpellary leaves separate not only at the shortened into the one-seeded indehiscent achene of the anemone or buttercup (fig. 2, e, f). In the achene of the grasses (which similarly represents the capsule of the ancestral lilies) the thin dry pericarp becomes inseparable from the seed-coat (hence the term caryopsis, fig. 2, c, d); in many trees (e.g. hazel) it becomes hardened and thickened as a nut. In composites (fig. 2, a, b), too, the achene is practically a nutlet, although often (on account of its being inferior) termed a cypsela. Less extremely reduced representatives of the various multicellular ovaries to which such fruits correspond are afforded us by borages or labiates, in which the two-celled ovary of the primitive solanaceous type becomes, as in thorn-apple, &c., subsequently divided into four parts: these (see fig. 2, i, h), however, are here so arrested as only to develop a single ovule in each loculus (of which the subsequent growth brings about the perplexing appearance of the 'gynobasic' style). The four ripe 'nutlets' into which the four-lobed ovary of these forms commonly breaks up were not unnaturally mistaken by the old botanists for naked seeds. In Umbelliferae we have another characteristic form of schizocarp, as all such fruits are termed which split up without truly carpellary dehiscence, although the tendency to this can be seen still to have some influence. Here the separate portions (or mericarps), each resembling an achene or nut, are two in number, and when ripe swing off upon the ends of a forked carpophore (fig. 2, j, k).
In exceptional cases we have the pod of some Leguminosæ and the silqua of some Crucifers—e.g. radish, snapping off into one-seeded joints, instead of dehiscing longitudinally in the regular way. This simply comes about where the swellings corresponding to the seeds become unusually large, leaving narrowings between them, and thus giving the pod a strength of form too great for the usual tension of ripeness to overcome (fig. 2, g). To confuse such distinct types of fruit under a common term (lomentum), and to separate them from the normally dehiscence capsules to which they really belong, and to place them among the purely 'schizocarpous' fruits we have been describing, although still too customary, are merely examples of the reasoned mistakes inseparable from a purely descriptive anatomy, but from which the evolutionary standpoint is at length delivering us.
So far all our fruits have been dry; but a new physiological 'principle of fruit-making' is necessary to comprehend those in which the pericarp is succulent. For, just as the effect of fertilisation is seen in many animals to extend beyond the mere ovum to the parent organism, and also in many of sides but also at the base, so curling inwards and projecting the seed. In Colchicum, white hellebore (Veratrum), and their allies (Melanthaceæ) the dehiscence is characteristically septicidal, the carpels separating instead of the loculi opening: the remaining majority of Liliaceæ are loculicidal. Where, however, the placentæ remain more or less completely upon a central column from which the valves are detached, the dehiscence is said to be septifragal.
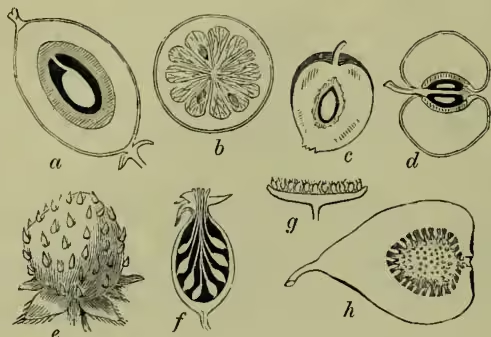
In henbane (fig. 1, c), Anagallis, &c. the dehiscence is circular (circumscissile); the possible explanation of this as a disarticulation of the united carpels by their leaf-bases is, however, rendered difficult through the separated portion being a mere lid. Many-celled capsules are numerous in which the leaf-opening or dehiscence is greatly reduced from completeness, witness the valvular and porous dehiscence of the Lychnis and of the poppy (fig. 1, a) respectively. Such cases clearly point us to those of carpels which do not open at all. Such indehiscent fruits, produced from carpels so persistently embryonic, are, as we might expect, usually short, few or one-ovuled, and, for the most part, little specialised. Thus the follicle of the Ranunculaceæ of more specialised floral character becomes the lowest plants, so it is in the case before us. Even in fruits which are dry on ripening we have seen that the ovaries or loculi, on which no demand is made for the growth of fertilised ovules, become reduced or disappear. Sometimes it may be merely the coats of the seed (as in the pomegranate) which undergo the complex histological and chemical changes which we sum up as those of succulence and ripening; at other times largely their placentas, as in the gooseberry and currant. Yet, as in these, the innermost tissue of the ovary may become succulent as well. In the orange also the familiar succulent tissue in which the seeds are immersed are the enlarged succulent cells of the endocarp; the grape too gives a characteristic example of soft endocarp. These may all be classed as berries or baccate fruits, for the distinction of the succulent product of an inferior ovary as a berry, from that of a superior one, as a uva or grape, need hardly be allowed to increase our nomenclature. A pepo is merely a berry in which the epicarp is thick and tough (e.g. a melon, with which the orange and pomegranate may be reckoned). Where the succulent change, instead of primarily affecting the deeper tissues of the fruit, and so producing a berry, leaves the endocarp hard, we have evidently a well-contrasted type—the drupaceous or stone-fruit. The endocarp here forms a more or less complete ‘stone’ around the kernel or seed, the difference from an ordinary nut being due to the succulence of an outer layer, as mesocarp, with a more or less leathery outer skin, the epicarp. The plum, peach, and nectarine are the most obvious examples; but, since we may have many carpels thus transformed, we may have an aggregate fruit or syncarp of tiny drupes. The walnut and even cocoa-nut are hence not true nuts (see NUT). The immature succulent mesocarp of the former is familiar in pickles, the walnut we crack being merely the stony endocarp (which is exceptionally specialised in being set free by the bursting of the mesocarp on ripening). The familiar cocoa-nut fibre is the fibro-vascular tissue of the mesocarp, the fruit being thus broadly comparable to a peach which has wizened while still young and stringy. But, as in the kindred grass, the coats of the ovule further unite to the endocarp.
The numerous carpels of the strawberry, although, of course, corresponding to those of the allied raspberry, remain mere nuts; here, however, the subjacent portion of the floral axis or receptacle becomes succulent. In the perigynous or epigynous Rosaceae the same change may take place; hence the rose-hip is a succulent axis, enclosing a multitude of nuts. The apple or ‘pome’ is more akin to the drupe, since the carpels, here deeply sunk in the upgrown floral axis, develop a hard endocarp corresponding to the stone of a drupe.
Fertilisation may even be followed by succulent or other thickening of the floral envelopes, or of the floral axis with subjacent bracts—the various cupules, as of acorn, beech, hazel-nut, &c., being of this nature. Or we may have a spurious fruit developed at the expense of an entire inflorescence, as in the pine-apple, Dorstenia, and fig. See INFLORESCENCE.
Fruits Important to Man.—The list of the fruits of any importance is much shorter than would at first be supposed, as may be seen from the following enumeration (practically that of Frank), which distinguishes those native or cultivated in northern Europe (Germany and Britain) from the more important foreign fruits, and of course employs the terms stone-fruits, berries, &c. in their merely popular sense.
(1) Indigenous or Cultivated: (1) Apples or Pip-fruits.—Apple (Pyrus Malus), Pear (P. communis), Medlar (Mespilus germanica), Quince (Cydonia japonica), Service-berries (Sorbus torminalis and S. domestica), to which may be added Hips (Rosa canina, &c.) and Hawks (Cratagus Oxyacantha), also Cornel-berries (Cornus mascula).
(2) Stone-fruit.—Peach and Nectarine (Persica vulgaris), Apricot (Prunus armeniaca), Plum (P. instititia), Cherry (P. Cerasus and P. avium), Damson (P. domestica), Greengage (P. italica), Sloe (P. spinosa), Cherry-plum (P. cerasifera), &c.
(3) ‘Berrics.’—Grape (Vitis vinifera), Strawberry (Fragaria vesca, elatior, &c.), Raspberry (Rubus Idaeus), Bramble or Blackberry (R. fruticosus), Gooseberry (Ribes Grossularia), Red Currant (R. rubrum), Black Currant (R. nigrum), Barberry (Berberis vulgaris), Black Mulberry (Morus nigra), White Mulberry (M. alba), Bilberry or Blaeberry (Vaccinium Myrtillus) with its minor congeners, Juniper (Juniperus communis).
(4) Nuts or Shell-fruit.—Hazel-nut (Corylus Avellana), Filbert (C. tubulosa), Walnut (Juglans regia). See NUT.
II. More Important Fruits of Warm, Temperate, and Tropical Regions: (1) Stone-fruit.—Date (Phoenix dactylifera), Olive (Olea europaea), Mango (Mangifera indica), Tahiti-apple (Spondias dulcis), Mombin Plum of West Indies (S. Mombin), Avocado Pear (Persea gratissima), Iaco or Cocoa Plum (Chrysobalanus iaco), Sapota Apple (Achras Sapota).
(2) Berries and Berry-like Fruit (in widest sense of succulence).—Banana and Plantain (Musa paradisiaca), Pine-apple (Ananassa sativa), Fig (Ficus Carica), Bread-fruit (Artocarpus incisa and integrifolia), Custard Apple (Anona squamosa, &c.), Baobab (Adansonia digitata), Orange, Lemon, Lime, Citron, Shaddock, Pompelmoose, Forbidden Fruit, Bergamot, and other species of Citrus, Pomegranate (Punica granatum), Guava (Psidium piriferum), Rose-apple of East Indies (Jambosa domestica and vulgaris), Tamarind (Tamarindus indica), Carob or Locust Bean (Ceratonia siliqua), Papaw (Carica Papaya), Pumpkin (Cucurbita Pepo, &c.), Melon (Cucumis Melo), Water-melon (C. Citrullus), Cucumber (C. sativus), Tomato (Lycopersicum esculentum), Lotus (Diospyros lotus), Jujube (Zizyphus vulgaris), Mangosteen (Garcinia Mangostana), Prickly Pear (Opuntia vulgaris).
(3) Nuts or Shell-fruit.—Cocoa-nut (Cocos nucifera), Almond (Amigdalus communis), Chestnut (Castanea vesca), Litchi or Lee-chee (Nephelium Litchi) (really, however, a shelled drupe), Brazil-nut (Bertholletia excelsa), &c.
Chemical Composition of Fruits.—Our knowledge of the chemistry of fruit may be dated from the analyses of Fresenius (1857). But because of the innumerable varieties of almost every cultivated fruit, the effects of different soils and climates upon these, and still more of the fluctuation due to better or worse seasons, the results of any one chemical analysis would tend to convey an idea of undue precision. Thus—e.g. while the ratio of sugar to free acid in certain grapes of an ordinary wine-year was found to be 16 to 1, in a very bad year it sank to 12, and in a very good year rose to 24. Hence a broad outline may be of more general use than the statistics of any one analysis.
The percentage of water may be taken as varying from 78 to 80 in the grape and cherry, as from 82 to 85 in plums, peaches, apples, and pears, as 82 to 87 in brambles, currants, &c., and as much as 95 in the water-melon. The proportion of insoluble residue—skin and cellulose, stone and seed—obviously also varies greatly with succulence and ripeness, but may be taken, one fruit with another, at not less than from 4 to 6 per cent. Unripe fruits may contain a notable proportion of starch, but this is fermented on ripening into glucose and other sugars, fruit-sugar, grape-sugar, cane-sugar, or (in Sorbus) sorbin. The only fruits which retain starch in important quantity are those of the banana, bread-fruit tree, and baobab; hence the exceptional nutritive value of these. The olive alone yields a notable proportion of oil. The proportion of sugars varies exceedingly, dates, dry figs (48 per cent.), and raisins (56 per cent.), again very important foods, heading the list. Grapes of course stand high, from 12 to 18, indeed sometimes as much as 26 per cent., cherries from 8 to 13, apples 6 to 8, pears 7 to 8, plums 6, red currants 4.75, greengage 3.5, peach and apricot only 1.5. The proportion of pectin bodies is, however, exceedingly notable, especially in fruits such as the three last named. In unripe fruits (as also in roots) we find pectose, a body apparently related to cellulose, but easily transmuted by a natural ferment or by boiling with dilute acid into pectin, , and its allies. These are all more or less soluble in water, with which they readily form a jelly (whence the peculiar consistency of our fruit-preserves). The proportion of soluble pectin and gum varies considerably and is of great importance to the blandness and agreeableness of fruit, the harder and more common apples having considerably less than 3 per cent. and the best rennets nearly 8. The harsh red currant, indeed, like berries in general, has exceedingly little (0.25 per cent.); while the apricot has as much as 9, the greengage 12, and the peach 16—a circumstance which explains the peculiarly melting quality of these fruits, especially the last named. The free acid also varies greatly, from 2.4 per cent. in the red currant, 1.4 in the raspberry, and nearly as much in the sourest cherries, to 0.5 in sweet cherries and a minimum of 0.1 or less in the sweetest pears. That of apples and of grapes, of course, varies greatly, but both may generally be taken at from 1 to 0.75, while the apricot and peach stand at 0.3 or 0.4. The acid is primarily malic, but citric, acetic, oxalic, tannic, and others may also be present.
The quantity of albuminoids is of course small, in fact inadequate to render most fruits a staple food. Yet it is by no means inappreciable, ranging from nearly 5 per cent. in the majority of fruits to 7 or 8 in the grape (2.7 in raisins), and above 1 in the melon and tomato. Hence to acquire albuminoids equal to those of one egg we must eat 1½ lb. of grapes, 2 lb. strawberries, 2½ lb. apples, or 4 lb. pears. To replace 1 lb. starch = 5½ lb. potatoes, we need 5.4 lb. grapes, 6.7 of cherries or apples, or 12.3 of strawberries (see FOOD).
The quality of fruits depends largely upon the proportion of sugar, gum, and pectin to free acid, largely also upon the proportion of soluble to insol- uble matters, but in very great measure also upon the aroma. This quality is due to the presence of characteristic ethers, often accompanied by essential oils, although not of course in ponderable percentage. Cultivation and selection operate strongly on all three factors.
Keeping of Fruit.—Many of the finest fruits undergo very speedy decomposition, which, as distinguished from the intrinsic processes of ripening, is due to the attacks of bacteria, moulds, or yeasts; and the problem of their preservation is therefore primarily one of preventing these. In damp and stagnant air, especially with considerable or frequent changes of temperature, these fungus pests multiply with special readiness; hence a fruit-room must be cool and shady, yet dry and airy, and the fruit carefully gathered rather before full ripeness, handled so as to avoid in any way bruising or tearing the skin, and laid out and occasionally looked over so that rottenness in one may not affect the rest. Under these conditions apples especially may be kept for many months; indeed many varieties of fruit—e.g. winter-pears—require these conditions for satisfactory ripening. On antiseptic principles we see how it is that the dense-skinned and wax-coated grape can be so largely imported in sawdust, or how unripe gooseberries, and even very perishable pears can be kept for months similarly packed in well-sealed jars in a cool place. The process of preserving with sugar in jars promptly covered up is similarly an antiseptic one; but in the systematic application of antiseptic principles we may still look for considerable progress in the preservation and transport of fresh fruit upon a large scale. The method of drying fruit has also been in use from remote times, especially with dates, figs, and raisins.
Of late years more attention has been bestowed on fruit-growing in Britain, and a large area of land is devoted to fruit-culture. But difficulties in distribution and the cost of transport have not infrequently the effect of glutting the available markets in good years, and making prices wholly unremunerative. By far the most of the fruit grown in Britain is produced in the counties nearest London. On the other hand, the reduction in the cost of ocean transit has largely increased our imports of fresh as well as dried fruit, all of which are free of duty, save figs, fig-cake, plums preserved otherwise than in sugar, prunes, and raisins (on which the duty is 7s. per cwt.), and currants (at 2s. per cwt.). The annual imports of fresh oranges and lemons have a value of over £2,000,000 per annum; of currants and raisins, £1,600,000; and of other fruit, over £2,000,000.
In the United States, the extension of the fruit-growing area has been very great; orange-growing in Florida and some other southern states is now a great industry; and in California (besides wine-producing), the preparation of rearing and growing of oranges, figs, and other fruits is carried on on a large scale. Then the United States imports annually—largely from the West Indies—fruits and nuts (especially bananas and cocoa-nuts) to the value of 17,000,000 a year, while exporting fruits and nuts to the value of 5,000,000. Fruit is cultivated in the Himalayas for Anglo-Indian use, and apples grow magnificently there.
See GARDENING, ORCHARD; the articles on APPLES, PEARS, PEACHES, and the various fruits; WINE, CIDER, &c.; PRESERVED PROVISIONS; and works on fruit-culture by Cheal (new ed. 1892), Thomson (1881), Fish (1882), Burbidge (1881), Du Breuil (1886), Hogg (1885), Wood (1880); and for America, by Downing (1876), Fuller (1881), Roe (1886), and Thomas (1876).