Flower. The observations of childhood, like the popular language in which these so largely tend to survive, are apt to deny flowers altogether to plants in which these are not conspicuous. Yet the unmistakable succession of the flower by the fruit and seed compels us in some measure to argue back from the presence of one or other of these to the previous existence of flowers, even where they may not usually have been noticed, as, for instance, in the oak. The observant walks of spring and early summer soon convince us that not one even of the soberest forest-trees but has its blossoming time, while no one can escape noticing in mid-summer the abundant blooms of the grasses and nettles. Horticultural experience, too, convinces us that the overpowering majority of plants brought from other lands flower sooner or later if the right conditions can be insured them, and thus we gradually reach the generalisation that among the plants which exhibit stems and leaves, none save the mosses, the ferns and horse-tails, the club-mosses and selaginellas, with a very few other inconspicuous and unimportant forms, fail to agree in the production of flower and seed. We have here evidently a broad principle of classification, and if our knowledge of the reproductive function of the flower has meantime developed so far as to repeat the discovery of its definite sexuality, we shall feel the appropriateness of Linnæus's general term of Evident-flowering or Phanerogamia (q.v.), yet also approve the mingled caution and speculative which made him apply that of Cryptogamia (q.v.) to the latter, since it was reasonable to suppose that the reproductive process (Gr. gamos, 'wedlock') evident (Gr. phaneros, 'well-exposed') might in the latter be concealed (Gr. cryptos).
The examination of so many flowers, or even of a few representative ones, will have shown us that the central organs in which the seed arises exhibit considerable variety alike in number and detail. Thus, distinguishing the essential 'seed-box' as the ovary, we find that there are many simple separate ones in the buttercup or the rose, while the apparently single ovary of the apple or the tulip turns out on section to be five- or three-celled respectively, each cell containing the future seeds or ovules. We find, too, above the ovary a surface usually viscid or hairy, to which pollen readily adheres, and this we call the stigma, noting also that while sometimes practically sessile upon the ovary, as in buttercup or poppy, this is oftener raised upon a column, the style.
Encircling the seed organs we find the stamens, their stalks or filaments bearing heads or anthers. We see the latter shedding forth a yellow dust, the pollen, the same as that which we have noted upon the stigma of some of the flowers. This suggests one or two simple experiments. If we cut off the stigma, or cover it so that pollen cannot reach it, the incipient seeds or ovules will never mature into seeds proper capable of germination. Conversely, if we remove the stamens of this and other flowers, so that the same isolation of the stigmas from pollen comes to pass, the same failure of the seed follows. Whereas, when pollination of any particular stigma is effected (whether by the natural agency of insects or the wind, or more convincingly, by the actual help of the experimenter), seed forthwith matures in the corresponding ovary. And if the pollen be from a plant exhibiting some characteristic varietal difference, the resulting seed, when grown in its turn, will show the influences of this 'crossing' in a more or less marked degree (see HYBRID, HEREDITY). In this way the conception of sex in flowers, of immemorial date as respects those of the Date-palm (q.v.), has been extended in quite modern times. The carpels and stamens being thus the 'essential organs,' we recognise as merely 'accessory organs' the parts lying external to these, despite their frequent conspicuousness and beauty. In both monocotyledons and dicotyledons we have usually two circles or whorls of these parts, but in the former they are usually very similar, and so are usually spoken of merely as inner and outer perianth-whorls respectively, and their parts as perianth-segments. In most dicotyledons the corresponding whorls are known as corolla and calyx, since the colour and magnificence of the blossom usually depend upon the former (Lat. corolla), while the latter is commonly reduced to a mere verdant cup (Gr. calyx). The parts of the corolla or petals, and those of the calyx (sepals) are hence usually recognisable at a glance, as in the buttercup or rose. Yet the corolla may be reduced to inconspicuousness, as in the Christmas rose or globe-flower, or even vanish altogether; while the calyx may assume splendours which would be noteworthy even for a corolla, witness the marsh-marigold or garden clematis. Petaloid characters may be even assumed by Bracts (q.v.). Sepals, and still more frequently petals, may become more or less completely united, as so familiarly in the common primrose. In monocotyledons both whorls of the perianth may unite, as in the lily of the valley. Peculiar specialisations of form also abound, especially in the corolla, witness pansy or balsam, dead nettle or snapdragon. The calyx, too, may have its peculiarities, as in wallflower or poppy. The very modes of their arrangement in the bud differ widely (see ÆSTIVATION); while the position and arrangement of the separate flowers opens up a new field of study (see INFLORESCENCE).
Applying to the essential parts corresponding collective names, we have then innermost of all the pistil or gynæcium (its separate or constituent members being termed carpels); next the androecium of stamens. We have thus four whorls of organs, each presenting apparently independent and often wide differences in different flowers; hence we understand how in the attempt conscientiously to take note of all this multiplicity of floral detail the nomenclature of descriptive botany almost inevitably became so intricate and cumbersome (see BOTANY) as to obscure and retard that very progress towards the recognition of the order and unity of the floral world which it was the aim of the whole science to reach. Keeping this aim, however, in view, we are compelled to rise from the description of separate plants to the problem of classifying them into groups (species, genera, orders, classes, &c.) according to the degree of their common resemblance. It soon becomes evident, as it did to the earliest systematists, that it is mainly upon the flowers that our classification must depend. And having seen the multifariousness of these, we are in a position to appreciate the aid of the Linnean or 'Artificial System' of orders and classes, when to ascertain or record all that is known of a new plant our primary task is but the simple one of noting the number of its carpels and stamens. We comprehend better, too, the reluctance with which the claim of the 'Natural System'—that classification must depend upon the sum of like characters—was conceded, since this principle leads us back to unravel all the perplexing intricacies of which the device of Linnæus so summarily cut the knots.
A principle of fundamental importance for the natural system was however reached, and descriptive botany made a vast step, from the empirical to the rational level, as the conception of the morphology of the flower (see MORPHOLOGY) became gradually established. Sepals and petals, stamens and carpels thus lost their utter distinctness, and fell into a single category with the leaves as the appendages of an axis; their differences thus becoming understood as specialisations for their respective physiological purposes, in no way inconsistent with fundamental unity of structure and development. The simplest observer soon realises that a sepal is a sessile leaf, and a stamen a stalked one, and only needs to dissect a double rose or a white water-lily to convince himself by the discovery of a perfect series of transitional forms between petal and stamen of the essential unity of these. The spiral instead of whorled arrangement which we find in some simpler floral types (Ranunculaceæ, &c.) is thus immediately intelligible as a transition to the ordinary phyllotaxis of leaves. But the botanist has hardly yet exhausted the more refined applications of this principle; thus, since a leaf may not only have stalk and blade, but side pieces or stipules, we have frequently to take note of these in the flower, just as in the analogous case of leaves reduced as bud-scales (see BUD). Nor does the utmost detail fail us: thus, since the stipules of opposite leaves, although normally separate (nettle), constantly tend to unite (wood-geranium) or may even develop like the ordinary leaves (bedstraw), so it happens in many flowers. What the descriptive botanist empirically termed the outer calyx or epicalyx of a strawberry or lady's mantle is thus rationalised as the united stipules of adjacent sepals; and similarly the perplexing whorl of external yet younger stamens in the Geraniums (q.v.) and a few other types is easily interpreted as similarly united stipular developments of the ordinary whorl of stamens. By help of the leaf, in short, we are learning more and more completely to interpret and unify the peculiarities of floral structure.
Returning to simpler cases, the union of sepals and petals not only recalls that of opposite leaves, but anticipates what we exceptionally find among stamens, as in the filaments of the mallows, or the anthers of the composites; or among the carpels in a majority of orders. That in the types with united corolla such union of the inner parts should also be more frequent and thorough than in those of separate parts is not to be wondered at; hence, for instance, the clubbed stigma of the lily of the valley as compared with the distinctly lobed—i.e. more separate stigma of the tulip.
Another great series of structural differences becomes cleared up when we bear in mind that appendages, however crowded and united, must arise upon an axis, however shortened. One or more internodes of the floral axis are, indeed, occasionally well developed, as in a lychnis or a passion-flower: but, generally speaking, their growth is checked at a quite embryonic stage. Since the early days of the natural system great attention has been paid to the position and origin of the floral parts. In the lily or the buttercup the stamens are seen to arise directly under the gynoecium, and are accordingly described as hypogynous; in the strawberry they arise in a circle at about the same level as the bases of the young carpels, and are consequently termed perigynous; while in the iris or the rose they seem perched upon the very top of the ovary, which is hence described as inferior. From the distinctness of the floral axis (then termed thalamus), the orders of hypogynous dicotyledons with separate parts became grouped as Thalamifloræ, while those with perigynous or epigynous stamens and separate parts were termed Calycefloræ, from the apparent origin of the stamens upon what was commonly regarded as a united calyx. All this, however, becomes intelligible when we combine the study of vertical sections of the adult flowers with the facts of their development. In the simplest hypogynous type we see the whorls of floral appendages standing in regular succession upon the conical floral axis just as they developed; the internodes having all developed equally. In the perigynous type, however, we see that the growth of the upper portion of the floral axis has been outstripped by that of its lower and as it were more vegetative region, which hence grows up around it; the epigynous type thus being only the extreme case of this, just as in inflorescences the spike becomes flattened into a capitulum, and this finally hollowed at the apex, or rather raised at the circumference into a cup, or even flask, as in the case of the fig. See INFLORESCENCE.
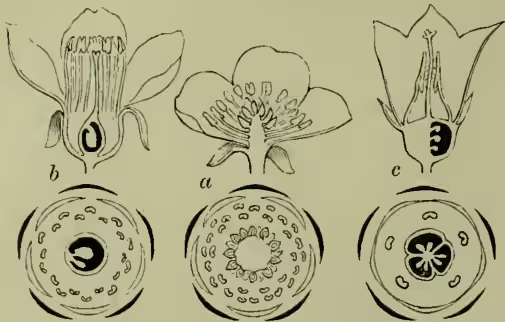
The large body of dicotyledonous orders in which the petals were united were grouped as Corollifloræ, and finally the small and apparently degenerate types in which calyx, corolla, or both were absent were termed Incompleteæ. The convenience of this system is so great that it is still retained in the Genera Plantarum of Bentham and Hooker. Its importance, but also its dangers may be well seen by comparing fig. 1, in which figures of types of as many well-marked Thalamifloral, Calycefloral, and Corollifloral orders are paralleled within the single order Liliaceæ (fig. 2), when with these we reckon the closely allied Amaryllids. In the same way the wide contrast between the usually splendid Calycefloral or Corollifloral and the inconspicuous Incomplete type might be paralleled within the range of a single order, witness rose and lady's mantle, or lilac and ash (Oleaceæ).
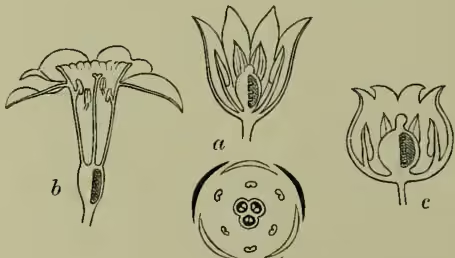
Though thus losing an absolute principle of classification, we gain a conception of simplicity underlying floral differences. From the mere empirical description of flowers as we find them we begin to penetrate the rationale of their development, to comprehend something of what we may call the principles of flower-making. Starting again from a simple and tolerably central type like that of the lily, we find among its immediate allies types here of increasing floral magnitude, there of diminished splendour and size; witness tulip and lily of the valley. We see how the more magnificent types tend to be always few or single-flowered, while the smaller and more modest types bear flowers in abundance; how the former are more individualised examples of the ordinal type, while the latter seem more embryonic. The one type tends to be richly coloured, often indeed complexly marked; the other tends to be paler, even white or positively greenish. When departures from regularity of form occur it is the more splendid forms which become bilaterally symmetrical; and conversely it is among the more embryonic types that the petals or other parts become united. The more vegetative habit of the lily of the valley as compared with that of the tulip is here of interest, since we can correlate this obvious constitutional difference with the facts already outlined, or with such an otherwise incomprehensible detail as the production of many ovules by the tulip, but of few by the more vegetative and consequently less reproductive blossom of the lily of the valley. Even were such facts comparatively isolated they would be suggestive; but a wider examination of the monocotyledons brings us face to face with the existence of the same contrast in groups of all extent. Those extreme types, as it would seem, at once of floral differentiation and of vegetative habit, which we know as grasses and orchids, are alike clearly referable to the simple liliaceous ground-plan of structure (see the floral diagrams in fig. 3), and the question hence arises—must we not regard these simply as extreme variations, greater in degree, but of no different kind than those which we have recognised among the Liliaceæ proper? An examination of the orchids or the grasses themselves, still better of the arums, yields a similar result: the floral orchids have their degenerate vegetative types, small, many-flowered, few-seeded, it may be even green; while even the typically crowded and degenerate florets of the arums may recover, as we see by viewing the whole inflorescence, no small measure of floral magnificence. Passing to the dicotyledons we find the same result in reviewing them orders by order, often indeed genus by genus (e.g. Clematis, Thalictrum, Delphinium, and Senecio). We thus comprehend the otherwise perplexing fact that of all supposed natural alliances it is that of the Incompletæ which has most broken down, their 'orders' having long been recognised as the degenerate (i.e. vegetative) representatives of many widely distinct orders. The bearing of these considerations upon our theory of variation and consequently upon our general conception of evolution cannot here be entered upon; it must suffice if the existence of order and simplicity amid the infinitude of floral detail has been rendered more apparent.
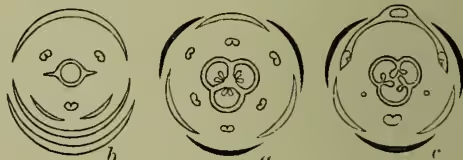 Grasses (q.v.). (c) Specialised modification usual in Orchids (q.v.)." loading="lazy">
Grasses (q.v.). (c) Specialised modification usual in Orchids (q.v.)." loading="lazy">The problems just now raised will be found more fully discussed under VARIATION, &c.; while the classification of flowering plants is more naturally treated under VEGETABLE KINGDOM. The structure of the stamen, with the development of its pollen, are naturally described under STAMEN, and the process of fertilisation of the ovule under OVULE; while that deepest interpretation of the flower which compels us to view the phanerogam as not only the most evolved of the cryptogams, but really the most profoundly cryptogamic of them all, is necessarily given under GYMNOSPERMS, since a knowledge of these in relation to the higher cryptogams is indispensable. It remains, however, to glance briefly at the flower from the physiological side, in its adaptations to fertilisation.
Fertilisation of the Flower.—From the frequent separation of the sexes in flowers borne upon the same individual (monœcism—e.g. Begonia), or upon different individuals (diœcism—e.g. nettle, dog-mercury, red or white campion), it is evident that pollen must somehow be borne from male to female flowers. The artificial assistance of the wind-fertilised Date-palm (q.v.) has been already referred to as indicating some comprehension of the process from immemorial times. The hermaphroditism of most flowers, however, long misled botanists into thinking of the flower as normally fertilised by its own pollen; and so obstinate was this error that even the publication of Sprengel's Secret of Nature Discovered, with its wealth of careful observation even supported by accurate figures, failed to dispel it (see BOTANY); and this admirable book fell into oblivion until its rehabilitation by Darwin, followed by Delpino, Hildebrand, Fritz Müller, and many subsequent investigators.
Just as the familiar spores of ferns or horse-tail are scattered by the wind, so it is with the pollen of such primitive flowers as those of Conifers and Cycads (see GYMNOSPERMS). In the cryptogams, however, germination of the spores takes place upon the ground apart from the parent plant altogether; but in the flower, since the female spore or embryosac remains retained within its sporangium or ovule, and this in turn in monocotyledons and dicotyledons (angiosperms) within a carpellary leaf which usually does not even open until the maturation of the fruit (see FRUIT), it is manifest that only those male spores (pollen grains) can be effective which are conveyed to the receptive surface (stigma) of the carpellary leaf, and there germinate until they succeed in penetrating as far as the ovule and embryosac. Hence the utility of wind or insects for the transport of pollen. In this regard it is evident that the help of spore-eating insects might early have been of service, since spores might readily thus be conveyed adhering to their bodies. It has been already noted (see CHLOROPHYLL) that the colouring matter of flowers, like that of vernal and autumnal leaves, must be associated with phases in the constructive or destructive changes of chlorophyll. Since the reproductive process especially checks those of vegetation, we have here an agency for the production of floral colour, which, by rendering the reproductive shoots more conspicuous to insects, would be constantly aided by natural selection. Upon the latter process, indeed, the customary explanation of the origin of floral colour and markings solely depends. An analogous advantage for the attraction of insects would be given by the overflow as 'nectar' of any excess of the sugary sap so largely used up by the flower itself, or similarly by the disengagement of perfume. A constant adaptation between flower and insect being in such ways established, further specialisations arise. Thus, while in many flowers—e.g. crucifers, the stamens and stigmas are ripe simultaneously, a 'want of time-keeping' is frequently observed, the stamens becoming protandrous—i.e. ripening before the stigmas, as typically in Geranium (q.v.), so that self-fertilisation is impossible, and a physiological separation of the sexes (dichogamy) is thus insured. Or where the ripening remains simultaneous, two or even three forms of flower (dimorphism, trimorphism) may occur in different individuals of the same species, so rendering cross-fertilisation by insects indispensable (see PRIMROSE, LOOSESTRIFE). The individual flower may also become peculiarly specialised: thus, the nectary may become so deep as to be accessible only to insects with long probosces, as bees to the exclusion of flies, butterflies and moths to the exclusion even of bees, it may be even to some particular species only. Birds, too, may replace insects (see HUMMING-BIRD, HONEY-BIRD). Mechanical adjustments are also to be found in many of the more specialised types, witness the stamens of the sage, which are rocked forward by the bee on entering the flower so as to dust his back with pollen just where it will be rubbed off upon the stigma of the next flower. The most extraordinary variety and complexity is, however, that presented among the Orchids (q.v.).
How some flowers (e.g. Stapelia, Rafflesia) attract the services of flesh-flies by the odour and even colour of carrion; how others like Arum and Aristolochia may entrap and detain the fertilising insect until well dusted with pollen, are examples which can but be mentioned. Nor can we describe the defences of flowers from rain or from ants, &c. (see, however, CATCHFLY, HEATH, &c.).
In wind-fertilised plants the flowers are usually comparatively small and inconspicuous, but numerous and closely aggregated, often in spikes, heads, or catkins; the floral envelopes are usually small and greenish; one or both whorls are indeed frequently absent (Incompletæ). The stamens are few but often versatile as in grasses, the ovary reduced, commonly only one-seeded, but with one or more styles and stigmas, the latter often exuberantly branched or feathery, so catching the small, smooth pollen. The stigmas are ready before the pollen is shed (protogynous dichogamy); and flowering often takes place as in many forest-trees before the development of the leaves. The pollen grains may be lightened with air-sacs as in the pine, may be helped by gravitation as in maize or bulrush, where the male inflorescence grows higher than the female, or may be scattered explosively as in the nettle or artillery plant. The utility of all these special characters of wind-fertilised flowers is thus no less obvious than that of the peculiarities of those dependent upon the visits of insects, and the natural selectionist is hence accustomed to draw from both of these remarkable sets of adaptations many of his strongest arguments for the development of even the most complex organic structures through the cumulative selection of minute spontaneous varieties (see DARWINIAN THEORY). Some facts, however, such as the repeated development of the characters of wind-fertilised flowers in quite unrelated types, seem rather to indicate the possibility of a constitutional interpretation similar to that indicated on the previous page (see VARIATION).
It is thus manifest that the flowers of every species in nature thus invite and reward an observant interest far other than that of the mere collector, systematist, or even morphologist. Further information will sometimes be found under individual headings—e.g. CAPRIFICATION, GERANIUM, VALLISNERIA, &c.; these cannot, however, exhaust the special works upon the subject.
See, for general reference and full bibliography, Müller's Fertilisation of Flowers (trans. by Thompson, Lond. 1882); also Kerner's Pflanzenleben, Bd. ii. (Leip. 1889); and Hooker's Students' Flora (1888). Among popular introductions to the subject those of Lubbock and Grant Allen ('Nature' series, &c.) are well known.