Respiration, or BREATHING, is a part of the life of all organisms, animal and vegetable. It is a series of chemical changes, the first of which is the absorption of oxygen into the body, and the last of which is the excretion of carbonic acid. The association of this intake of oxygen and excretion of carbonic acid with the same organs, the lungs, is due to the fact that both the food-stuff and the waste-stuff are gases, and not to any immediate connection between them. Necessarily any organ adapted to the diffusion of a gas from the air into the blood must also be adapted for the diffusion of a gas from the blood into the air; that is, supposing that the living membrane, of which the lung essentially consists, which separates the air from the blood, acts, so far as the diffusion of gases is concerned, as a dead membrane; even if it has any effect arising from the fact of its being a living membrane it is probable that it will behave in a similar way to both the ingoing and outgoing gases. We shall first describe the process as it occurs in man and other Mammalia, and then give a sketch of comparative respiration, the various methods adopted in other types of animals for effecting the same purpose. The respiration of plants is dealt with in the article on VEGETABLE PHYSIOLOGY. For the general relation of the function of respiration to the other bodily functions, see PHYSIOLOGY.
In all animals in which the parts of the body are nourished by the circulation of a stream of food material, the blood, there must always be two distinct sets of processes to consider: (1) the maintenance of the blood in a normal state, by the supply of fresh food-matter from time to time, and by the elimination of waste-matter; (2) the nutrition of the individual tissues and cells of the body by the blood-stream (see CIRCULATION). Applying this to the function of respiration, we shall have to consider (1) the manner in which oxygen is supplied to the blood and carbonic acid gas removed from it; (2) the manner in which the cells are able to take oxygen out of the blood and cast into it their useless carbonic acid; also the changes that take place within the cells between the intake of the oxygen and the output of the carbonic acid. These two sets of processes are usually described as the outer and the inner, or tissue, respiration. Some writers include in the term outer respiration the absorption of oxygen by the cells from the blood, and the excretion of carbonic acid into the blood, and restrict the term inner respiration to the actual changes that take place within the cells. It is evident that outer respiration corresponds to the processes of digestion and absorption to which food materials other than gaseous are subjected. It must be evident, too, that the mass of our present information refers to outer respiration; the changes that take place within the cells, of which inner respiration is a part, are the whole mystery of the objective side of life.
OUTER RESPIRATION.—In all animals which possess a blood-stream the outer respiration is carried on by the simple diffusion of oxygen into and of carbonic acid out of the blood through a thin membrane from and into the air or water in which the creature lives. As already noted, it is possible that the fact of this membrane being a living one may in some ways modify the otherwise simple processes of diffusion. The essential structure, therefore, of all breathing organs, lungs, gills, or tracheæ, must be the same: a thin membrane exposed on the one side to the oxygen-containing medium, air or water, in which the animal lives, on the other side to the blood flowing in a network of thin-walled vessels, so that the gases that have to pass in and out of the blood are only separated from the air or water from which and into which they have to pass by thin partitions—by the membranous wall of the breathing organ, and by the thin wall of the blood-vessels. Animals such as the frog, which have thin skins, can breathe with—i.e. the gases can diffuse through—the whole surface of their bodies if the under skin is well supplied with blood-vessels. A frog for this reason can live for days without its lungs, but if its skin be rendered impervious to gases will die very quickly, even with the lungs intact. But, as all the Mammalia have thick skins, this method of breathing must be reduced to a minimum, if indeed it exists at all.
In outer respiration we have two things to consider: (1) The manner in which fresh supplies of oxygen are pumped into the lungs, while the poisonous carbonic acid gas is pumped out. This may be called the mechanics of respiration. (2) The manner in which oxygen passes from the air in the lungs into the blood, and is held in the blood, and the manner in which the carbonic acid in the blood passes out of the blood into the air-chambers of the lungs. This may be called the chemistry of respiration.
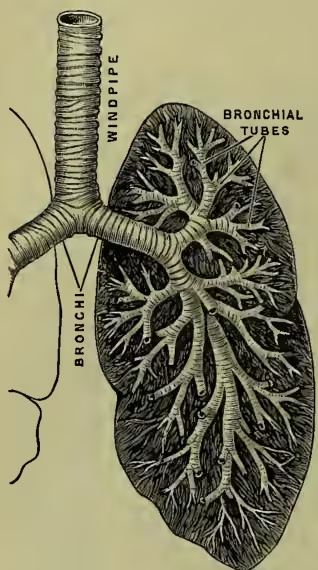
Structure of Respiratory Mechanism.—This mechanism consists of the lungs, a series of minute air-chambers with a network of capillaries in the walls, the air-passages from the air-chambers of the lungs to the outer air, and the chest-walls with their muscles, which act like bellows and change the air in the lungs. The essentials of structure that a lung must possess have already been emphasised. The simplest lung that we can imagine would be an elastic membranous bag, well supplied with blood-vessels, and with a pipe connecting it with the air; the most complicated that exist are essentially of that construction, the complications that occur having for their object merely the enlarging of the surface exposed to the air. Let us begin with the air-passages. There are first the nose and mouth; these join the upper part of the gullet, known as the pharynx (see illustration at DIGESTION). From the pharynx arises the windpipe (trachea); this passes through the voice-box (larynx) into the chest-cavity; there it divides into two passages (the bronchi); the bronchi go on dividing again and again, generally into two; the ultimate divisions (the bronchioles) open into clusters of air-chambers. The air-chambers are about th inch in diameter. It has been estimated that there are some 725 millions of them, and that their total surface is about 2000 square feet. The walls of the air-chambers are formed of a thin membrane in which the blood and lymph capillaries ramify. Minute openings lead from the air-chambers into the lymph spaces of the membrane. The membranous walls are partly formed of elastic tissue. It is this that gives to the lungs their elasticity. The larger air-passages (trachea and bronchi) are kept open by horseshoe-shaped plates of cartilage; muscles stretch between the poles of the horseshoe, complete the ring, and permit the size of the passages to vary, at the same time resisting over-distension when the internal pressure rises. These larger air-passages are lined by a mucous membrane, containing mucous glands; the innermost layer is a ciliated epithelium; the cilia lash upwards, and thus keep the passages free from mucus and remove foreign particles. As the passages become smaller they lose their cartilages, and the muscles form a continuous circular layer. The lungs are invested by a membrane (the visceral pleura). At the root of the lungs this membrane is continuous with a membrane which lines the chest-cavity (the parietal pleura). The space between the two is the pleural cavity; it is in reality a large lymph space, and communicates with the lymphatics of the pleura. Owing to the air-pressure within the lungs the two pleurae are closely pressed together, the lungs entirely filling the chest-cavity. If the chest-wall be punctured the lungs partially collapse owing to their elasticity, and the respiratory movements are unable to move the air in the lungs.
The chest is an air-tight chamber enclosing the lungs and the heart. The walls of the chest are formed of bones (the ribs, sternum, and backbone) and muscles; the bones and muscles are so arranged that the size of the chest-cavity can be altered. In this way the chest acts as a bellows and moves air in and out of the lungs. The ribs are sloped slightly downwards, especially after an expiration; when an inspiration is taken certain muscles fix the upper ribs, and those muscles connecting the ribs to each other contract and the ribs are raised, and thus the size of the chest-cavity is increased. At the same time a flat muscle called the Diaphragm (q.v.), which separates the chest-cavity from the rest of the body-cavity, and which after an expiration is arched upwards (by the pressure of the abdominal viscera upon it, the viscera in turn being pressed upon by the abdominal walls), forcibly contracts, becomes flatter, and therefore enlarges the size of the chest-cavity, forcing the abdominal viscera downwards and causing the abdomen to protrude. (The relation of the lungs to the other main organs will be seen in the illustration at ABDOMEN; see also that at DIAPHRAGM.) In these two ways, then, the size of the chest-cavity may be increased. The result of this enlargement is that the pressure of the air within the cavities of the lungs is lowered; air therefore from without rushes through the nostrils (one ought not to breathe through one's mouth) down the windpipe into the lungs, and thus a fresh supply of oxygen is introduced. The movements which produce this result are known as the inspiratory movements. In making an expiration the reverse effects are produced; the chest-cavity is made smaller, the pressure of the air in the lungs increases, and some rushes out through the nostrils into the air until the pressures inside and outside are equalised. An ordinary expiration is effected by the elasticity of the lungs, by the fall of the ribs, unsupported by the contraction of the muscles that caused an inspiratory movement, by the elasticity of the cartilages of the ribs which were twisted during inspiration, and by the elasticity of the abdominal wall which was forced outwards by those viscera pushed downwards by the diaphragm. An ordinary inspiration is therefore the result of a number of active muscular contractions, while an ordinary expiration is the result of mere passive elasticity of the parts concerned. There are certain other respiratory movements to be considered. During inspiration and expiration the glottis (the opening between the vocal chords of the larynx; see the illustration at LARYNX) undergoes a rhythmical widening and narrowing; this movement is greater in forced than in quiet breathing. And during inspiration the nostrils dilate; in most cases perhaps the inspiration has to be rather a forced one before they do so. Forced respiration occurs when the supply of oxygen is insufficient, or when carbonic acid accumulates in the blood. Any muscle that can aid in enlarging and decreasing the size of the chest-cavity is called into play. The average amount of air, in the case of an individual 5 feet 8 inches in height, that goes in and out of the lungs at each inspiration and expiration is about 20 cubic inches; this is called the tidal air. By means of forced inspiratory movements the ingoing tide may be increased by 120 cubic inches; by means of a forced expiration the outgoing tidal air may be increased by 90 cubic inches. After the most forced expiration possible there always remain within the lungs about 90 cubic inches of air. So that if we take as deep a breath as possible, and then make as forced an expiration as we can, we shall drive out cubic inches of air. This is termed the respiratory capacity. Since the tidal air is only 20 cubic inches, and 180 cubic inches remain in the chest after an ordinary expiration, it follows the air directly changed during respiration is not that really within the lungs themselves, but is that within the nose, windpipe, and larger bronchi, the pipes that result from the branching of the windpipe. Therefore the changes of the air within the essential parts of the lungs are the result of diffusion between it and the purer air of the bronchi, aided by the rush with which the tidal air flows in.
The ordinary respiratory movements differ in the two sexes and at different periods of life. In young children the chest is altered in size chiefly by the movements of the diaphragm, and the protrusion of the abdominal wall during inspiration is therefore very marked. In men also it is the diaphragm which is chiefly operative, but the ribs are also moved. In women it is the movement of the ribs, especially the upper ones, which is the most extensive. The respiratory rhythm is the relation of the acts of inspiration and expiration to each other as regards time. It may be expressed as follows: In. = 3, Ex. = 4, pause = 3. The number of respirations in a healthy person is about fourteen or eighteen per minute; it is greater (nearly double) in childhood. It varies according to circumstances, exercise, rest, health, disease, &c.; in disease it may fall as low as seven or rise to a hundred.
The proportion of respiratory movements to heart-beats is about one to four, or one to five; in health they vary together. Since the heart and the lungs are contained in the same air-tight cavity, it follows that the variations in size of the heart as it beats must rhythmically affect the pressure of the air in the lungs, causing a succession of minute puffs of air to leave and enter the nostrils. Similarly the alterations in pressure within the chest-cavity affect the heart. Increase of pressure or expiration must (owing to the arrangement of the valves) help the blood to flow out of the heart. Decrease of pressure or inspiration must, for the same reason, help the flow of blood into the heart. The pressure which the expiratory muscles, aided by the elasticity of the parts concerned, can exert is on the average equal to that of 4 inches of mercury. The inspiratory muscles can lower the pressure within the chest-cavity by a pressure equal to that of about 3 inches of mercury below that of the atmosphere; the greater part of the energy of the inspiratory movements is used in overcoming the elasticity of the lungs, chest-walls, and abdominal walls. The respiratory sounds are two in number: (1) the tubular sound, heard over the windpipe and the larger bronchi, probably due to friction of air in these passages; (2) the vesicular sound, heard over the whole chest during inspiration, probably caused by the sudden dilation of the small air-chambers of the lungs, and to friction in the smaller passages. During a quiet expiration there may be no sound; when present it is very soft and indistinct, probably due to the air passing out of the air-chambers.
The Nervous Mechanism of the Respiratory Movements.—Although all the muscles concerned in the movements of breathing are voluntary muscles—i.e. can be made to contract by an act of will—yet respiration is normally an entirely involuntary act. This is obvious from the fact that during sleep, or during absence of consciousness caused in any way, respiration goes on as well as during wakefulness. Further, although we may at will breathe or cease to breathe, yet we cannot by any effort of the will suspend the respiratory movements for longer than at most a few minutes at a time. We have seen how many are the muscular movements involved in breathing, and it is obvious that the adjustment as to time and intensity of contraction of all these muscles must be a very nice one—in technical phrase, they must be co-ordinated. Such co-ordination must always be the result of a nervous mechanism, and this co-ordination, together with the fact of the rhythmical nature of the respiratory movements, suggests that the whole must be under the dominance of a nervous centre. The position of this centre has been ascertained by experiment; the whole of the upper part of the brain may be removed, and yet breathing will be unimpaired; but if a certain part of the medulla (see BRAIN, and illustration, Vol. II. p. 388) be injured or removed then all respiratory movements cease at once; the centre must therefore be in that part of the medulla. The centre is bilateral, for destruction of one-half of the medulla is followed by paralysis of the respiratory muscles of that side only. Further, we must conclude that, since inspiration is in its muscular movements antagonistic to expiration, there is an inspiratory centre and an expiratory centre in each of the two halves of the respiratory centre; but, as already noted, the expiratory centre is active only in forced respiration. The similar centres on each side are so co-ordinated that they act as one centre. This compound centre then is to be regarded as regulating the respiratory movements. We have said that if the medulla be injured the respiratory movements cease at once, and that from this it is concluded that the respiratory centre is in the medulla; but in young animals it seems that the movements may continue after destruction of the medulla, or may be produced by the reflex stimulation of some centre by irritating the skin. This subsidiary centre must be in the spinal cord; but it almost certainly is a subsidiary centre, though the matter is not quite settled yet.
Now is the centre 'automatic' in its discharges of nervous impulses, or is it reflexly stimulated into action by the arrival of stimuli from some other part of the body? We know by ordinary experience that the centre may be influenced from without, by impulses arising from higher parts of the brain, as when by will we alter the respiratory rhythm, or when it is affected by emotions, and also by impulses arising from the stimulation of sensory surfaces, as when cold water is dashed against the skin. It is found by experiment that the centre may be influenced in two distinct ways: (1) by nervous impulses; (2) by changes in the blood.
Nervous impulses may affect either the inspiratory or the expiratory part of the centre. It seems that all afferent nerves—i.e. nerves in which the impulses travel towards and not away from the central nervous system—may influence the respiratory centre (see NERVOUS SYSTEM). But the vagi (nerves that are distributed to all the viscera) seem to be in specially close relation, beginning as they do close to the respiratory centre in the medulla, and ending in the lungs. If one vagus be cut there is not much effect upon the breathing; but if both are cut then the breathing becomes slower and deeper. If the end nearest the centre of one of them be stimulated the respiratory rhythm is generally quickened; by a certain strength of stimulus it may be made normal; if the strength of the stimulus be further increased the inspiratory movements may be made before expiration is finished; this effect increased to a certain extent must obviously result in a standstill of all respiratory movements; the chest-walls remain in the inspiratory place. But occasionally it happens that stimulation of the central end of a vagus, after both have been cut, produces a further slowing of the movements—they may indeed be entirely stopped; in this case the chest-walls remain in the expiratory phase. From these results it is concluded that the vagus contains two kinds of fibres that affect the respiratory centre, one kind that increases the respiratory movements, another that inhibits them; and, further, that when one kind is active in causing increased inspiratory movements the other kind is active in causing depressed expiratory movements. Further, if air be drawn out of the lungs, thus imitating expiration, an inspiratory effort is made; if air be forced into the lungs, thus imitating an inspiratory movement, an expiratory effort is made. Therefore we may conclude that expiration stimulates the inspiratory centre, and that inspiration stimulates the expiratory centre. That the effects from which these conclusions are drawn are due to the stimulation of the vagus endings in the lungs is shown by the fact that they do not occur when the vagi have been divided; and that they are not due to alteration in the state of the essential gases of the blood is shown by the fact that they may be produced by forcing an indifferent gas, such as nitrogen, in and out of the lungs. The respiratory pump is therefore a self-regulating mechanism.
If we cut the vagi the respiratory rhythm usually becomes slower, and the movements are deeper; therefore normally stimuli are constantly passing up the vagi to the centre, and accelerating the discharge of impulses by the centre. Still, an accelerating effect is not the same thing as an initiating stimulus. Further, since respiration goes on when the higher parts of the brain are removed, impulses proceeding from above are not essential; and since when the spinal cord is cut below the medulla the movements of the nostrils and vocal chords continue (although of course all others cease), the centre works independently of sensory impulses arriving from any nerve, except the cranial nerves; and since these cranial nerves may be divided, if the medulla and spinal cord be left intact, without any effect upon the respiratory movements, we may conclude that the centre is automatic in its action, but may be influenced from without.
The more venous the blood the greater is the activity of the centre; when the blood reaches a certain state of impurity convulsions arise. We may conclude that the state of the blood affects the centre directly, and not reflexly, by stimulating the endings of afferent nerves in various parts of the body; because if the supply of blood be cut off from the medulla alone the same effects are produced. Venous blood differs from arterial blood in containing less oxygen and more carbonic acid. The deficiency of oxygen is the cause of the greater activity of the centre, because if an animal breathe an atmosphere of nitrogen the carbonic acid does not accumulate in the blood, and yet convulsions occur; whereas if the animal breathe an atmosphere containing sufficient oxygen but excess of carbonic acid, then the convulsions do not occur, but the animal may become unconscious through some of the higher centres being poisoned. When in action the centre discharges motor impulses down various nerves to all the muscles concerned in the respiratory movements. If any of the nerves be cut the movements of the muscles supplied of course cease, since they are no longer stimulated by impulses proceeding from the centre.
The Chemistry of Respiration.—We have now to explain the passage of oxygen from the air-chambers of the lungs into the blood that circulates in the vessels of the chamber-walls, and the passage of carbonic acid from the blood into the air within the lungs.
In order to understand what follows we shall have to study the laws of diffusion (see also DIFFUSION). A gas consists of a great number of separate molecules moving with great speed. The number of these molecules in a cubic inch of a gas (at ordinary temperature and pressure) is estimated about or 1,000,000,000,000,000,000,000. Each molecule is so small that the space between adjacent molecules is large compared with the size of the molecules; therefore, each molecule during its movement has a large path free from collision with other molecules. The average speed of a molecule varies with the temperature, increasing as the temperature rises. The molecules lying near the surface of any mass of gas will constantly impinge upon the boundaries; these impacts are so numerous and so close that they produce an apparently continuous pressure all over the boundary. This pressure obviously depends only upon the density (number of molecules in unit-space) and the temperature (average speed of the molecules) of the gas. Further, the molecules of a gas are so far apart that when two or more gases are mixed their molecules interfere so little with each other that each gas exerts the same pressure upon the walls of the containing vessel as it would do were it alone present. In such a case the total pressure is the sum of the two or more partial pressures of the several gases. If the space in which a gas is enclosed be diminished the molecules are brought nearer to each other, until a point is reached at which many of the molecules apparently act upon each other in such a way as to become more complex molecules, thus forming a liquid in the lower part of the vessel with its gas in the upper part. The complex molecules are still in motion, and interchange, or diffusion, constantly takes place between the two regions. The number of molecules leaving the gaseous region depends only upon the state (temperature and density) of the gas. The number leaving the liquid depends only upon the state (temperature and density) of the liquid. When the diffusion takes place in a closed space a state of equilibrium of interchange is soon reached. In the lungs the liquid molecules of the oxygen of the blood are being constantly moved past the common surface between the air and the blood; the inflow therefore of oxygen from the air into the blood is greater than the outflow from the blood to the air. On the other hand, the gaseous carbonic acid in the air is constantly removed from the common surface between it and the blood; and therefore the outflow of carbonic acid from the blood into the air is greater than the inflow from the air into the blood. This picture of the state of matters that regulate the interchange of gases in respiration is simpler than the reality. The further complexity will be described immediately.
We must know what are the laws governing the diffusion when the gas above the liquid is not the gas of the liquid, as is the case when air rests upon a surface of water. Some of the molecules of the air will become entangled in the liquid, will form the liquid of the particular gases within the other liquid, and then the state of affairs will be as before, so far as the gases, and their liquids, of the air are concerned, and a state of equilibrium between each of these gases and its own liquid will be formed. But now suppose that the liquid and the gas have a special chemical affinity for one another, as is the case with the oxygen of the air and a substance in the blood, and as is the case with the carbonic acid of the blood and a substance or substances in the blood. As soon as the gas has diffused into the liquid the chemical compound will be formed; but now the reverse effect will begin, dissociation of the compound will occur, but slowly, because a greater violence of collision is necessary. Therefore, other things being equal, less pressure will be needed to maintain equilibrium, because fewer liquid molecules of the compound will become gaseous, and therefore fewer gaseous molecules need become liquid to preserve equilibrium. Indeed, it is found that at a certain temperature and a certain pressure the dissociation scarcely takes place at all; but if temperature be raised, or if the pressure be lowered to a certain point, then the dissociation will be very rapid.
These laws of diffusion apply to the gases of the blood. In the investigation of these gases a sample of blood is placed under the receiver of an air-pump (thus imitating, though exaggerating, the normal pumping action of the chest-walls), the gases extracted are passed through various solutions which retain the several gases, and thus they may be estimated and examined. The quantity of oxygen obtained from arterial blood is greater than that obtained from venous blood. The arterial blood of a dog yields for every 100 vols. at ordinary pressure and C. 58.3 vols. of mixed gases when the external pressure is reduced to zero. This mixture is composed of 23.2 vols. of oxygen, 34.3 vols. of carbonic acid, and 1.8 vols. of nitrogen.
If blood took up as much of these gases by mere diffusion as water does, it would contain 0.86 vols. of oxygen, 1.2 vols. of carbonic acid, and 1.6 vols. of nitrogen. Therefore it is evident that, while the nitrogen is merely diffused into the blood, the oxygen and the carbonic acid must be combined with some substance or substances in the blood. If we gradually lower the external pressure of the atmosphere upon the blood we notice that at any given temperature (at which the combination can exist) the pressure may be lowered to a certain point without much gas coming off, and that at that point the gases begin to come off rapidly. This is another proof that the gases are combined and not merely absorbed in the blood; for in case of simple absorption the gases come off in equal amounts for equal lowerings of pressure. The amount of the gases that can be taken from blood-plasma (free from blood-cells) is 0.26 vols. of oxygen, 35.26 vols. of carbonic acid, and 2.24 vols. of nitrogen. The great mass of the oxygen is, therefore, not in the plasma, but in the corpuscles; while the great mass of the carbonic acid is in the plasma. The oxygen is found to be united to the red colouring matter, of which the red blood-cells are chiefly composed. This substance is called hæmoglobin. It is not so easy to determine in what combination the carbonic acid exists in the plasma. A certain amount is found in the red corpuscles (though the above figures do not show it); indeed, some writers consider that the hæmoglobin of these cells is the chief carrier of carbonic acid. The effect of lowered pressure upon blood-plasma, so far as regards carbonic acid, is much the same as it is upon solutions of sodium hydrogen carbonate. Some writers believe that the carbonic acid exists in the plasma in the form of sodium bicarbonate. Others believe that it may be in the form of bisodium hydrogen phosphate. The presence of red blood-corpuscles has a very marked effect upon the disengagement of carbonic acid under lowered pressure; it hastens it considerably. This effect appears to be due to the presence of oxyhæmoglobin.
The total pressure of the atmosphere is 760 mm. of mercury. The partial pressure of oxygen in the air is 159.6; of carbonic acid, practically zero; of nitrogen, 600.4. Oxygen does not leave arterial blood until the partial pressure falls to 29.64, nor venous blood until the pressure falls to 22.04; these therefore are the partial pressures of oxygen in arterial and venous blood. Carbonic acid does not leave arterial blood until the partial pressure falls to 21.18, and venous blood until it falls to 41.04. Therefore blood exposed to air would readily gain oxygen and lose carbonic acid. But the air in the part of the lungs where the respiratory interchange takes place is not the same as the air surrounding the body; the partial pressures of expired air will be nearer the true numbers; they are—of oxygen, 121.6; of carbonic acid, 33.4; of nitrogen, 600. But even expired air is not the same as air within the alveoli; for the air taken in and out of the lungs (tidal air) only enters and leaves the larger respiratory passages near the opening into the outer air; from these it diffuses into the air of the alveoli. The partial pressures of this air have been estimated by introducing a collector into the alveoli and taking out samples. Specimens of air collected in this way have been found to have the following partial pressures: Oxygen, 27.44; carbonic acid, 27.06; nitrogen, 705.5. It is difficult to believe that this is a correct estimate, for the difference between the partial pressure in the alveoli and that in the expired air is so enormous. However, assuming it to be correct, the following diagram will show the direction in which diffusion must take place.
| Venous Blood. | Alveolar Air. | |
|---|---|---|
| Oxygen ..... | 22.04 | 27.44 |
| Carbonic Acid..... | 41.04 | 21.04 |
The vertical line represents the alveolar and capillary wall; the arrows show the direction in which the gas molecules must diffuse. But if we compare the partial pressures in venous blood, in arterial blood, and in alveolar air, a very remarkable fact appears.
| Alveolar Air. | Venous Blood. | Arterial Blood. | |
|---|---|---|---|
| Oxygen ..... | 27.44 | 22.04 | 29.64 |
| Carbonic Acid.. | 27.06 | 41.04 | 21.04 |
The venous blood flows through the lungs, and issues as arterial blood, and yet the partial pressure of oxygen in arterial blood is higher than it is in alveolar air, the place from which it must have come; while the pressure of carbonic acid in arterial blood is lower than it is in alveolar air, the place to which it has passed. We must therefore conclude that the living alveolar wall has exercised some influence upon the gases in virtue of its secreting and excreting activity; it has done work against the molecular energies that produce diffusion. But the numbers given by various authors for the partial pressures of the gases in the various places differ, so that perhaps no thoroughly reliable conclusion can be drawn from them. Still in any case the slight differences of partial pressure, especially of oxygen, render the validity of any explanation of the rapidity of gaseous interchange within the lungs in terms of ordinary diffusion extremely doubtful. A possible aid to the interchange has recently been suggested in the sudden stroke of the heart, which would have an accelerating effect upon the liberation of gases from a fluid under low partial pressure; just as a tap upon the sides of a glass containing soda-water will cause bubbles of carbonic acid to be given off. Further, as already stated, some carbonic acid is combined with hæmoglobin. This combination is, like oxyhæmoglobin, dependent upon the partial pressure of the carbonic acid, and is easily given off when that pressure is lowered. Possibly the hæmoglobin may be an important carbonic acid carrier in the blood.
Effects on Respiration of the Quality and Quantity of the Gases of the Atmosphere.—The respiratory mechanism, as well as the whole body, is adapted to work with air of a certain composition, and at a certain pressure. The mechanism can adapt itself, within certain limits, to variations of composition and pressure. We have to state what these limits are, and what happens when they are overstepped. We shall study first of all, because of its practical importance, the results of breathing in a confined space, or in one insufficiently ventilated. The effect upon the air of course is that the proportion of oxygen is lowered, and that of carbonic acid increased. The first effect upon a person experiencing such a state of affairs is that a sense of mental and muscular fatigue occurs when the proportion of carbonic acid rises to 0.1 per cent., the normal proportion being 0.04 per cent.; and this is not due to the carbonic acid, but to the presence of organic matter, derived probably from the clothes, of the amount of which the carbonic acid happens to be a measure; for if pure carbonic acid be introduced into the air of a room, until the proportion rises to 1 per cent., no disagreeable sensations are experienced in breathing it. If the proportion of oxygen be still further diminished, or if by shutting the trachea of an animal all supply of oxygen to its blood be cut off, the oxygen of the blood begins to be used up, and carbonic acid begins to accumulate, and asphyxia sets in. There are three stages of asphyxia. (1) The breathing becomes deeper and more rapid, the blood-pressure rising at the same time. (2) The respiratory movements continue to increase in force and rapidity, extra muscles are called into play, the expiratory movements are especially marked; then all the muscles that can possibly aid in expiration are used, the excitement spreads to nearly all the muscles of the body, and convulsions set in; these violent efforts exhaust the body. (3) A stage of quiet, due to exhaustion, occurs; a few long-drawn inspiratory gasps are made, these die out; the whole body is convulsively stretched out, and death intervenes. When the trachea of a dog is artificially closed these events run their course in from four to five minutes; the convulsions appear at the end of the first minute, and cease suddenly within the second minute. In drowning death is often hastened by the entrance of water into the lungs. The time at which death from drowning occurs varies with the state of the animal at the time of the occurrence. Young animals—e.g. a puppy—in which the respiratory changes are less active than in adults, may survive an immersion of fifty minutes; but a full-grown dog rarely recovers after having been minute under water. For man, see below, p. 667. By training, as in the case of divers, the respiratory centre may be accustomed to bear the scarcity of oxygen for much longer than it can normally.
We next consider the effects of changes in the partial pressures of the gases of the atmosphere, the total pressure remaining more or less unchanged. Lessened partial pressure of oxygen, as already noted, results in asphyxia. Increased partial pressure of oxygen results in the phenomenon known as apnæa. After several very deep inspirations the state known as apnæa occurs, and it is easy to hold the breath for a longer time than usual. The usual explanation of this has been that the oxygenation of the blood is so complete that there is enough to last some time, and the centre is not stimulated by its absence or by the presence of the reducing stuff in the blood. Later authorities regard the cessation of respiratory movements which occur when oxygen is rapidly forced into the lungs by rapidly succeeding respiratory movements as due to fatigue of the respiratory apparatus. Increased partial pressure of carbonic acid tends to the accumulation of carbonic acid in the blood, ultimately producing a state of narcosis without convulsions. Decreased partial pressure of carbonic acid results merely in the carbonic acid of the blood being able to leave the blood with greater readiness. Alterations in the partial pressure of nitrogen have no effect. Ozone, instead of making the blood more arterial, as one might expect, makes it more venous, and causes irritation of the respiratory passages. Carbonic oxide combines with the hæmoglobin with more avidity than oxygen; consequently it interferes with due respiration. Sulphuretted hydrogen, acting as a reducing agent, has ultimately the same effect. Nitrous oxide (laughing gas) produces narcosis, and is used as an anæsthetic. Some gases—hydrogen, marsh-gas, and other neutral gases—have no effect. Some—chlorine, ammonia, &c.—cause spasm of the glottis, and so cannot be breathed.
Another point to attend to is the effect of variations in external pressure, the proportional composition of the atmosphere remaining unchanged. Sudden and great diminution of pressure will cause fatal convulsions, due to the sudden liberation of bubbles of the gases of the blood within the vessels; these plug up the smaller vessels, and affect the working of the valves of the heart, and cause asphyxia. If the pressure be gradually diminished, as in ascending a mountain, no effect even at considerable heights is experienced beyond a feeling of 'distress' often accompanied by bleeding at the nose. This is due to a derangement of the vascular system, the walls being constructed to meet a certain external pressure. If only the respiratory interchange of gases were concerned, the total external pressure might be reduced from 760 mm. to 300 mm., corresponding to a partial pressure of oxygen of 76 mm., and to an altitude of 17,000 feet, before the combination of oxygen with hæmoglobin, at the temperature of the blood, would be seriously affected. In various parts of the world there are people living at an altitude of 11,000 feet. If the pressure be still further reduced asphyxia occurs, but it is not quite the same asphyxia as that which results from absence of oxygen; the characteristic convulsions are often absent, while a rapid onset of feebleness amounting almost to paralysis occurs. Increase of pressure up to a pressure of several atmospheres is followed only by symptoms of drowsiness, due probably to increased pressure upon the whole organism rather than to a direct derangement of respiration. At a pressure of fifteen atmospheres, which corresponds to a partial pressure of oxygen of three atmospheres, the animal dies of asphyxia with convulsions as though from a deficiency of oxygen. The production of carbonic acid is diminished with increase of pressure—i.e. the oxidations of the whole body are lessened. At a certain point these oxidations cease, and the animal dies. All living things are killed by a too great pressure of oxygen. The oxidations of some other substances—e.g. phosphorus—are analogous; at a certain pressure they will not burn.
The effect of variations in temperature must not be overlooked. By variations in temperature we mean of course variations in the temperature of the body and of the blood, and not merely variations in the temperature of the surrounding medium, for these have normally, in warm-blooded animals (the temperature of cold-blooded animals varies with that of the surrounding medium), no effect upon the temperature of the body, owing to the regulating mechanism afforded by the vessels of the skin and vaso-motor system (see CIRCULATION). The temperature of an Eskimo is nearly the same as that of an African; and in a Turkish bath the temperature only rises a very little. In cold-blooded animals the oxidative and chemical processes of the body decrease with a lowered temperature, and increase with increase of temperature; but the reverse is the case with warm-blooded animals, for the temperature of the body in an atmosphere of low temperature is partly kept up by increased oxidation; but in fever—i.e. when the temperature of the blood is actually raised—the chemical activity of the body of a warm-blooded animal rises. Such an animal dies when the temperature of its blood rises to 45° C. or 50° C., a mammal at 45° C., and a bird at 50° C. Death is due to the fact that when the temperature rises to this point the partial pressure of the oxygen of the air is no longer sufficient to maintain the combination of oxygen with hæmoglobin. Theoretically a higher temperature might be survived if the external partial pressure of oxygen were proportionally increased.
INNER OR TISSUE RESPIRATION.—We now come to the last and most interesting part of our subject—the manner in which the oxygen of the blood enters the tissues, the use made of this oxygen by the cells of the tissues finally resulting in the formation of carbonic acid, and the manner in which this carbonic acid leaves the tissues and enters the blood. The term 'inner respiration' is by some writers restricted to the interchange of the gases between the tissues and the blood; but it is usual and more convenient to include in that term what is known of the uses made of the gases by the cells. We have spoken with confidence of this respiratory action of all the cells of the body, but we must not forget that it has not always been believed in, and even now is doubted by some. The original theory was that the oxygen was used, and the carbonic acid formed, in the lungs only. This was disproved when it was shown that there is more oxygen and less carbonic acid in the blood coming from the lungs than in that going to them. Next it was, and still is by some, thought that the oxidations take place within the blood; the cells of the tissues were imagined as pouring oxidisable matters into the blood. Usually very little matter capable of taking oxygen away from a loose combination can be found in the blood, but in that of asphyxiated animals more of such matter was found; this was explained by supposing that in asphyxia the oxidisable excreta from the cells accumulated in the blood through insufficiency of oxygen; but it has recently been shown that this reducing stuff only exists in the red blood-cells—i.e. in the reduced hæmoglobin—while lymph, which we might expect to find rich in such matters, it being into the lymph that most of the excreta of the cells are poured, is totally devoid of it. Lastly, the supposition that the cells of the tissues use the oxygen directly is so much in harmony with all our present ideas of animal physiology and with the facts of comparative respiration (one-celled animals breathe, and plants breathe, and in these there is no circulating blood) and of embryology (the embryo mammal breathes though its blood-vessels are not connected directly with those of its mother) that one is disposed to believe it without further proof.
The mode of interchange of gases between the blood and the tissues must be the same as that with which we are already familiar—viz. the diffusion from a place of high partial pressure to one of lower partial pressure. The fact that a low partial pressure of oxygen is constantly maintained within the tissues is one of the phenomena that constitute the mystery of life. We have already seen that even in outer respiration the living cells of the essential membrane of the lungs may apparently do work against partial pressure, absorbing more oxygen and excreting more carbonic acid than the differences of pressure will account for; it is therefore extremely probable that a similar state of activity is characteristic of the cells of the other tissues. Taking the more obvious facts first, we know that with any weight of body—i.e. with a given amount of tissue to be supplied with oxygen—the amount of oxygen taken in and of carbonic acid excreted varies with the activity of the organism and with the amount of work that it is doing; it is greater in youth than in old age, in wakefulness than in sleep, during the activity of secreting glands than when these are at rest, during the performance of muscular work than in repose; in this case it is the excretion of carbonic acid rather than the intake of oxygen which is especially marked. This last peculiarity brings us face to face with a remarkable state of affairs. The partial pressure of oxygen within muscular tissue is always practically zero—i.e. however low the external pressure of oxygen may be, none will leave the muscle. The effect of this of course will be, so far as ordinary diffusion is concerned, that oxygen will always be leaving the blood and entering the tissues. This oxygen is in some way stored up within the muscle-cells, so that a muscle will work for a considerable time without any fresh supplies of oxygen, even in an atmosphere of nitrogen. This explains the fact noted above, that during muscular work the excretion of carbonic acid is in excess of the absorption of oxygen. A supply of oxygen, however, is necessary for the maintenance of the irritability of the muscle, which soon falls off without it, probably before the supply of stored oxygen used for the performance of its work has been exhausted. This is about all that is known of the chemical changes connected with respiration within a cell. The oxygen enters it by diffusion, possibly aided by some vital activity; the rapid storing away of the oxygen and consequent readiness to absorb more is in reality an example of such activity; the oxygen is made use of within the cell for maintaining its life, for producing heat, for producing rapid decompositions which supply the energy of muscular contraction; finally the carbonic acid leaves the cell and enters the blood, possibly aided in this process by some process other than a simple diffusion. The respiratory changes of other tissues are probably similar to those of muscle; within them, within the lymph that bathes them, and within their secretions there is practically no free oxygen, while the pressure of carbonic acid, owing to its constant production within the cells, is greater within the cells, their secretions, and the lymph that bathes them than it is in venous blood.
There is another fact about respiration which is still a puzzling matter, and, since it results from the changes within the cells, is likely to remain so for some time. All the food of a meal, or its equivalent, is in about six hours oxidised into carbonic acid, water, and urea. This is obvious without any elaborate calculations from the fact that we may eat every six hours and yet not gain in weight, while, apart from the indigestible parts of the food, which do not affect the problem, the chief matters that leave the body are those mentioned above. Yet food-stuffs outside the body are not affected by oxygen at the temperature of the body. Various suggestions as to the possible reason for this have been made; but, since the phenomenon is obviously dependent upon the vital processes of cells, suggestions in terms of the principles of ordinary chemistry cannot carry us far.
Further information regarding respiration will be found in the last editions of the text-books of physiology recommended at the end of the article upon that subject. The diseases of the respiratory organs are dealt with in separate articles, BRONCHITIS, CONSUMPTION, PLEURISY, PNEUMONIA, TUBERCLE, &c.
Historical.—Aristotle (384 B.C.) thought that the object of respiration was to cool the body. He observed that the warmer the animal the more rapid the breathing, and transposed cause and effect. Galen (131–203 A.D.) experimented upon the mechanics of respiration, and knew something of the nervous mechanism. He believed that 'soot' and water were excreted from the body by the lungs. Malpighi (1661) described the structure of the lungs. Van Helmont (1664) discovered carbonic acid; Black (1757) observed that carbonic acid is breathed out of the body. Priestley (1774) discovered oxygen. Lavoisier (1775) discovered nitrogen, found the composition of the air, and taught that the formation of carbonic acid and water resulted from the combustion that took place in the lungs. Vogel proved the existence of carbonic acid in the venous blood; Hoffmann found oxygen in arterial blood. Magnus extracted and analysed the gases of the blood in both states.
Comparative.—Most of the Protozoa, all the sponges and stinging animals, and many simple worm-types live in water, which washes their surface and saturates their substance, the oxygen dissolved in the water serving the same purpose as that mixed with the air. While many worms breathe simply through their skin, many of the aquatic forms have structures specialised for respiration—modifications of the legs or tentacles or vascular outgrowths of the body-wall. In Echinoderms respiration is effected by the tube-feet, and sometimes by hollow 'skin-gills' as well. The crustaceans usually breathe by gills or through the skin; in Peripatus, Myriopods, and insects air-tubes or tracheæ ramify throughout the body. Scorpions have plaited sacs or 'lung-books,' which many regard as modifications of tracheæ; and these are developed in spiders also, with or, rarely, without the addition of ordinary air-tubes. The king-crab has a unique arrangement, consisting of plaited sacs or 'gill-books,' adapted for breathing in water. Some molluscs breathe simply by the skin, others have external gills, most have gills sheltered by the mantle, and air-breathing forms like snails have a mantle-cavity which serves as a lung. In Balanoglossus there are numerous respiratory clefts opening from the pharynx to the exterior; Appendicularia and young Tunicates have a pair of these; in adult Tunicates the primitive clefts are replaced by numerous secondary slits on the wall of the pharynx, through which water drawn in by the mouth passes into an atrial or peribranchial chamber and thence to the exterior; the same is true of Amphioxus. Clefts from the wall of the pharynx to the exterior are, indeed, characteristic of vertebrates, but beyond amphibians they are transitory embryonic structures, never used for breathing. This loss of functional gill-clefts is associated partly with the development of an embryonic birth-robe known as the allantois, which secures the aeration of the embryo's blood, and partly with the transition from aquatic to terrestrial life. In the hagfish the nasal sac opens into the mouth; in fishes this is only true of the double-breathing Dipnoi; in all other vertebrates air passes through the nostrils in and out of the mouth and lungs. In the hag and lamprey there are purse-like gill-pockets, and the respiratory arrangements are otherwise peculiar. In fishes gill-filaments are borne on the skeletal arches separating the gill-clefts, and the blood-vessels spread out on the filaments are washed by currents of water. Young Elasmobranchs have at first external gills and afterwards the internal gills characteristic of all fishes. The Dipnoi have gills, but they also come to the surface and gulp air, using their air-bladder as a lung, and thus pointing the way to amphibians. For, while almost all amphibians have gills in their youth, all the adults are lung-breathers, though some retain their gills as well. Among higher vertebrates there are many peculiarities, such as the single lung of most serpents, the balloon-like air-sacs around the lungs of birds, and the adaptations of cetaceans as aquatic lung-breathers, but the essential characteristics of pulmonary respiration are the same in all. The hæmoglobin, so important in respiration, occurs first in Nemeriteans, and is present in some other worms, some Echinoderms, a few Arthropods, some molluscs, and in all vertebrates except the Tunicates, Amphioxus, and a few exceptional fishes. But though hæmoglobin is not present in most invertebrates, analogous pigments are common, especially one called hæmocytoyanin, which turns bluish when oxidised.
ARTIFICIAL RESPIRATION.—When death is imminent owing to a cessation of the natural respiration movements, it may sometimes be averted by an imitation of them carried on regularly for some time. Such a condition may occur in disease (e.g. asthma, epilepsy), though very rarely; it is most common in suffocation, either by drowning, choking, or strangulation, and is sometimes met with also in poisoning by noxious vapours (e.g. carbonic acid, carbonic oxide, coal-gas, chloroform, &c.).
In order that any method may have a chance of being successful it is of course necessary that the entrance of air into the lungs be not impeded, either by a piece of food or by water in the windpipe, or by the tongue falling back and closing the upper opening. A piece of food may sometimes be removed through the mouth by the finger; if this fails the windpipe should be opened (see TRACHEOTOMY). In those apparently drowned the body should first be laid on the face, with the head low, and the thorax and abdomen pressed upon in order to expel fluids which may have been drawn into the trachea and bronchial tubes. The tongue may need to be held forward; this may be done by an assistant, or an elastic band passed round the tongue and the chin will effect the object.
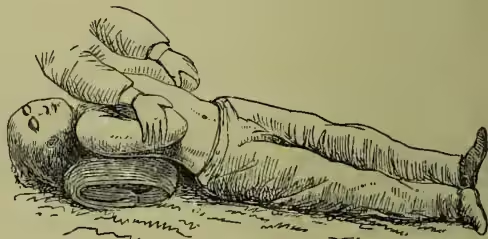
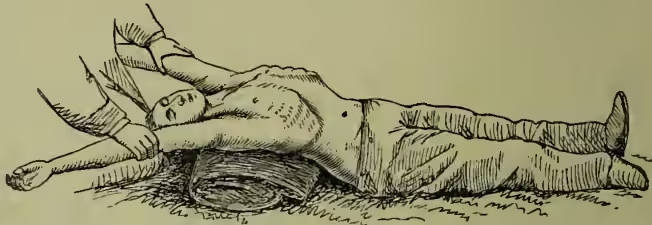
Numerous different methods have been devised for effecting the objects aimed at, and no general consensus of opinion has yet been arrived at as to which is the best. The methods fall into three divisions: (1) insufflation, or blowing of air into the lungs, either by the mouth or by means of bellows; (2) manual methods, in which external manipulations of the chest-walls are made to effect the entrance and exit of air; (3) electrical stimulation of the respiratory muscles. In all cases where artificial respiration is required every moment is of importance. It is doubtful whether life can ever be restored when the heart has ceased to beat for more than a few seconds; and when breathing has stopped failure of the heart's action is always imminent. That method is therefore best which can be applied with the least possible loss of time, so that under ordinary circumstances the methods which require bellows or electric batteries are out of the question. Direct insufflation, or blowing of air into the patient's lungs by the mouth applied to his mouth, is now hardly ever used except in the case of very young children. Of the manual methods those most in use are Marshall Hall's (1856), Silvester's (1857), and Howard's (1877). The second is certainly the most easy to learn, but is more fatiguing to carry out for a length of time than either of the others. In Marshall Hall's method the body is laid upon its face and rolled 'in what may be termed cradle fashion' from this position on to one side and a little beyond it (inspiration), and then back on to the face (expiration). In Silvester's method the patient is laid on his back on a plane, inclined a little from the feet upwards, and the shoulders are gently raised by a firm cushion placed under them, which also throws the head back. The operator then grasps the patient's arms just above the elbows, and raises them till they nearly meet above the head. This action initiates inspiration. The patient's arms are then turned down, and firmly pressed for a moment against the sides of the chest. A deep expiration is thus imitated. In Howard's method the patient is laid on his back with a cushion below the middle. The operator kneels astride his hips, places his hands with fingers spread outwards over the lower part of the chest-wall, and alternately bends forward, throwing his weight on the chest to imitate expiration, and springs back to allow the elastic recoil of the chest-wall to imitate inspiration.
Whatever method be adopted, the movements must be gently, regularly, and perseveringly carried on, at the rate of from ten to fifteen times in the minute; and when the faintest natural effort at respiration is observed they must at once be timed so as to reinforce and not to oppose it. In some cases life has been restored under artificial respiration when no respiratory movements have occurred for an hour or even several hours. In all cases, but especially in that of persons apparently drowned, artificial respiration should be conducted in a warm atmosphere, 90° F., or even more if possible, and should be supplemented by warmth applied to the body and by vigorous friction. In those apparently drowned recovery is very rare after complete immersion for five minutes or more. If stunning or fainting has occurred at the moment of immersion, so that the respiratory movements have been annulled or much diminished for the time, less water will have entered the lungs, and the chance of recovery may be greater. In other modes of death by suffocation, such as choking or strangulation, the action of the heart may continue longer, and restoration to life be therefore possible after a longer deprivation of air.
See the publications of the Royal Humane Society and Royal National Lifeboat Institution; various handbooks on ambulance work. A résumé and discussion of the various methods is given by Dr B. W. Richardson in the Asclepiad for 1890, p. 201.