Root. The exact use of the term in botany is not quite fixed. Old usage restricted it to the roots of seed plants, the organs of other plants which have the same general functions receiving various names; thus the roots of cryptogams were called rhizoids. The formal definition of root then used to be a part of a plant which produces no leaves, which has its growing-point protected by a root-cap, and which is developed endogenously—from the inner and not from the outer tissues. But it is more satisfactory to use the word in its physiological rather than in its strictly morphological sense, and to describe as roots all those parts of plants which (1) absorb nourishment from the soil or from water and not from the air, (2) fix the plant to the earth, (3) and grow, as a rule, downwards into the earth, and away from the light, not upwards, and towards the light, as 'shoots' do. Thus not only the complex roots of the seed plants, but also the rhizoids of cryptogams and the mycelia of fungi are called roots, in contradistinction to all those parts of plants which grow upwards towards the light, and which produce reproductive organs, which may conveniently be called shoots. But it is distinctly to be understood that organs of the same kind, from the morphological point of view, may have different forms and different functions; thus shoots may function as roots, and roots may grow up into the air as shoots. Further information as to the physiology of roots will be found in the article upon VEGETABLE PHYSIOLOGY; in this article it is chiefly the morphology of roots that will be dealt with.
As the leaf surface of a seed plant is developed the root system grows; in a large sunflower it occupies about one cubic yard, in a large tree hundreds of cubic yards. This system, resulting from the branching of the primary, secondary, and other roots, the ultimate divisions consisting of root-hairs, is so complete that scarcely the space of a quarter cubic inch is unoccupied. Abundance of water, to balance the transpiration, and of salts, for other purposes of plant-life, is thus secured. The root system of aquatic plants is small, water being easily obtained and transpiration slight.
All roots at first are thin hair-like organs; the greater thickness that those of the dicotyledons attain is due to a secondary growth. The thickened roots serve only as organs of attachment, and of course as conducting channels, but not at all as organs of absorption. These absorbing parts, root-hairs, are protuberances of the outer layer of cells of the roots. They arise shortly behind the growing tip, and they die off in a few days, so that only an inch or two of root bears root-hairs. In this way the root-hairs are continually brought into contact with portions of untouched soil.
A longitudinal section near the apex of a root of a seed plant shows the following arrangement of parts. Near the apex is the growing-point, a mass of actively dividing cells, called the primary meristem; by division these give rise to, below, several layers of simple cells (the root-cap), above, the main mass of the root, which, immediately above the primary meristem, consists merely of simple 'meristem' cells. A little higher up the meristem differentiates into three layers—the dermatogen, the periblem, and the plerome. These undergo further differentiation, so that a mature root consists of the following parts: (1) an outer layer, the epidermis, developed from the dermatogen; (2) several layers of simple cells, the cortex, developed from the periblem; (3) the fibro-vascular cylinder, an innermost mass of simple cells, and the pith, both developed from the plerome. The root-cap serves as a protection to the delicate growing-point of the root as it forces its way through the soil. It is continuously worn away by the particles of the soil, and as continuously reformed by the primary meristem cells. The epidermis binds the whole together, and many of its cells develop into long hairs, the absorbing root-hairs already mentioned. The cortex has no special function. The bundle sheath, when it occurs, separates the cortex sharply from the rest of the root. The fibro-vascular cylinder is the part of the root that acts as the channel for the ascending water and salts (crude sap), and gives it also its strength as an organ of attachment. The cylinder consists of smaller bundles of wood vessels and of bast fibres arranged alternately. The centre of a young root is occupied by pith.
Secondary thickening may occur in roots as in stems, and is due to the formation of a layer of actively dividing cells, the cambium. The bundles of wood and bast are placed alternately in a cylinder, and the cambium ring seen in transverse section waves in and out so that it passes inside the bast bundles and outside the wood bundles. The result of this is that the secondary thickening of roots results in a mass of tissue similar to that formed by secondary thickening of stems (see STEM). Thickened roots usually lose their original cortex and epidermis and gain a fresh covering in the following way. A layer of tissue, the root-sheath or endodermis, is differentiated which envelops the fibro-vascular cylinder; within this lies a layer of parenchyma, simple tissue; it is called the pericambium or phellogen. This layer gives rise to the new covering, which consists of two parts, phelloderm and periderm (which is of cork), corresponding to the original cortex and epidermis.
Forms of Roots of Seed Plants.—The primary root is merely the enlarged radicle of the seedling; it is the direct prolongation of the stem. All secondary roots arise from this first root; the secondary roots may give rise to others and so on until the common much-branched root is formed. If the primary root is very thin the whole system is called a fasciculated root; if the secondary fibres are also very fine it is called a fibrous root (e.g. the grasses); if the fibres are very much thickened it is said to be a tuberous root (e.g. the dahlia). When the primary root is much thicker than the secondary roots it is called a tap-root, and may be further classified according to its shape, as spindle-shaped (e.g. the carrot) or turnip-shaped. Much-thickened roots are usually storehouses of food material—e.g. the fasciculated root of the Turk's cap, lily, or the tap-root of the turnip. Roots are also described as fleshy (e.g. the beet-root), or as woody (e.g. the roots of trees). Fleshy roots usually die in the autumn; woody roots may live for many years, even when the shoots die down every autumn. Roots are usually buried in the soil, but they may be aerial, as for instance the roots of climbing plants such as ivy; these arise from the stem and fix the plants to their supports. In the tropics many plants have aerial roots; thus the mangrove forms forests in the swamps. The Indian fig has aerial roots from which, after they have become fixed in the ground, new plants may spring. Orchids are examples of the Epiphytes, which have aerial roots which merely fix the plants to trees but do not derive any nourishment from them. Aquatic plants often have roots which do not penetrate into the mud but float freely in the water. Many plants will form new roots after all the old ones have been cut off if they are placed in water or in a damp soil. This is taken advantage of by gardeners in their method of propagating such plants as fuchsias, oleanders, &c. from cuttings.
Esculent roots are numerous, and many roots also contain secretions either peculiar to themselves, or more abundant than in the other parts of the plant, and become therefore useful in medicine or in the arts, while some are very poisonous. The roots used for food, besides the Tubers (q.v.), Bulbs (q.v.), and Corms (q.v.), are generally those which are thick and fleshy. The plants to which they belong are of very different genera and orders—some of the natural order Cruciferae, as the turnip and others of the genus Brassica—some of the order Chenopodiaceae, as beet and mangold wurzel—some of the order Umbelliferae, as the carrot, parsnip, &c.—and some of the order Leguminosae.
Comparative.—Most of the Thallophytes (Algae, Fungi, and Lichens) have only a rudimentary differentiation into stem, leaf, and root, or none at all; but the most highly developed members of each division have an outward distinction of parts to which such terms may be given; still the roots even of these are distinguishable from the roots of vascular plants by the absence of a root-cap, and by their branching, which is never endogenous.
The Vascular Cryptogams—ferns, horsetails, lycopods, &c.—are highly differentiated in form and in their tissues. The roots arise in acropetal succession on the stem, or in many ferns on the petioles; their branching is monopodial or dichotomous; they are all alike—the first root never becomes like a tap-root, the lateral roots arise from the innermost layer of the cortex. The roots grow by successive divisions of an apical cell. The Selaginellae, a group of the Lycopodiaceae, are remarkable for having true roots, but a few have no root-caps on them. The growing-point too is formed by a primary meristem and not by an apical cell.
The roots of the Seed Plants, consisting of Gymnosperms—i.e. Cycadeae, Coniferae, Taxaceae, and Gnetaceae, and of Angiosperms—i.e. Monocotyledons and Dicotyledons, have already received a general description: a few points of comparison may here be repeated. The primary root is a continuation of the primary stem; it is directed towards the micropyle of the embryo-sac (see OVULE), and even it is of endogenous origin, its first rudiment being covered by the nearest cell of the suspensor. All the parts of the seed plants, shoots and leaves as well as roots, are distinguished from the parts of all other plants, except the Lycopodiaceae, and for a short time the embryos of some Coniferae, by having a small-celled primary meristem at the growing apices instead of the typical apical cell of many Algae, and of the Muscineae, Ferns, and Horsetails. The root-cap is formed, not as in the cryptogams, by transverse divisions of an apical cell, but by rapid division in the direction of the apex, and in gymnosperms from the periblem, in angiosperms from the dermatogen. Also the first rudiments of lateral roots, shoots, and leaves do not arise from single cells, as in the cryptogams, but from protuberances of a few small cells. The formation of a closed cambium ring in the primary and stronger lateral roots, and the subsequent secondary increase in thickness, is a characteristic of gymnosperms and dicotyledons, and does not occur in the cryptogams. This habit often results in the formation of persistent root systems, which in the monocotyledons are often replaced functionally by rhizomes, tubers, and bulbs.
The stems of the Cycadeæ are not unlike those of the Tree-ferns, but unlike them they have tap-roots; secondary roots appear above ground. In the Conifere the endosperm bursts the seed-coat at the root end, and the root, which develops a strong tap-root with lateral members, is thrust out.
The primary root of monocotyledons soon ceases to grow, lateral roots spring from the axis, each new root springs from a point higher up the stem, and being stronger than the older ones, there is no secondary thickening and no persistent root system. Some saprophytic monocotyledons form no roots at all. In dicotyledons an axis below the cotyledons is called the radicle, but the upper and often the larger portion consists of a part of the stem called the hypocotyl; the lower part is the true embryo root. The root is the first part to issue from the burst seed-coat; it grows and gives rise to lateral roots. If the primary stem grows vigorously the primary root also grows rapidly and forms a tap-root. If the growth ceases adventitious roots are often formed between the former lateral ones, which may themselves give rise to lateral roots. For further information, see Sachs's Physiology of Plants (1887) and Goebel's Classification and Morphology of Plants (1887).
ROOT-STOCK or RHIZOME is the name given to an underground stem when its shape is cylindrical; to a creeping stem, in short. It is easily distinguished from a root by the fact that it ends in a bud, and bears leaves or scales. It gives off roots every here and there. In the autumn the aerial leaves die down, but the rhizome lives through the winter, and in the spring its terminal bud goes on growing. Rhizomes are usually by the autumn well stored with reserve food-matter for the use of the bud in the following spring. Examples: the anemone, the lily of the valley, the yellow iris, many ferns.
Root, in Algebra, denotes any value of the unknown quantity in an equation which will render both sides of it identical (see EQUATIONS, INVOLUTION). The square root is that number which, multiplied by itself, produces the given number; the cube root, the number which, multiplied into itself and then into the product, produces the given cube; so with fourth root, fifth root, &c. The determination of the roots of equations, either formally or actually, constitutes the greater portion of the science of Algebra.
Root, GEORGE FREDERICK, an American composer, was born at Sheffield, Massachusetts, 30th August 1820, taught music in Boston and New York, studied a year in Paris, and returned to write numerous songs that became widely popular. Among them are 'Rosalie, the Prairie Flower,' 'There's Music in the Air,' and, during the civil war, 'The Battle-cry of Freedom,' 'Just before the Battle, Mother,' and 'Tramp, tramp, tramp, the Boys are marching.' His greater efforts, including a Te Deum, are less known. He died on the 6th of August 1895.
Root and Branch Men, a party in the Commons and out of it who supported a petition signed by 15,000 London citizens, praying that episcopacy might be destroyed 'root and branch.' Nathaniel Fiennes, Sir Harry Vane, and Hampden were of the party. A bill to give effect to the petition was read a first and a second time in 1641, but was ultimately dropped.
Root-parasites, plants which grow upon, and derive their nourishment from, the roots of other plants. Such are the Broom-ropes (Orobancheæ, q.v.), species of Thesium, &c., and the Rafflesias (q.v.), with other Rhizanthææ (q.v.).
Ropes. The staple fibre for ropes has long been Hemp (q.v.), but since the middle of the 19th century several other fibres have come largely into use. Hemp is better suited for cordage (the general term for ropes, cords, and twine of all kinds) than for weaving purposes, because it gives way when much folded into sharp folds more readily than other important textile fibres—flax, for example. Manilla hemp (Musa textilis; see ABACA), belonging to a quite different order of plants from the common hemp, makes the strongest ropes now manufactured, and is largely employed. Coir fibre, from the husk of the cocoanut, is another important rope-making material which, though long used in India, has in Europe only taken its place as a valuable cordage fibre in comparatively recent years. It is fairly strong, and, size for size, it has the advantage of being considerably lighter than either hemp or manilla. Unlike these, coir rope is never tarred for preservation. Sisal hemp, from the Agave sisalana of South America, though not nearly so strong as common hemp, is also much used, especially for ropes of small size. For certain purposes, such as driving parts of textile machinery, cotton ropes are largely employed. There are only two other rope fibres which as yet can be said to be of any importance in Britain—viz. the Sunn hemp (Crotalaria juncea) of India and the Phormium tenax of New Zealand. A large number of plants valuable for cordage grow in India, for a list of which, as well as for an account of the native process of rope-spinning, see Balfour's Cyclopædia of India.
As regards the strength of rope, it will be readily understood that in this respect specimens of the same kind of vegetable fibre will vary considerably. Healthy plants will yield a better fibre than those of more feeble growth, and some supplies of the same kind of material will have been more carefully prepared for spinning than others. The amount of care and skill bestowed upon the spinning process itself will also vary in the case of different manufacturers. Tarred ropes, again, though more durable, are not so strong as when left untarred; and as tar is liable to be impure it will happen that it weakens or injures the fibre more at one time than at another. These matters will partially explain why an unexpected difference not unfrequently occurs in the strength of two similar ropes which might be supposed to be equally strong. In the valuable work by W. G. Kirkaldy on his father's System of Mechanical Testing (1891) he gives the breaking stress of a large number of tested ropes, and in his remarks on this section of his experiments points out that the strength of some of these is much lower than it ought to be. He thinks that this can only be accounted for by adulteration of the fibre. It is probable that jute is sometimes mixed with better material. In one instance he found some rubbish called 'batch' hidden inside the rope. The following examples will show approximately the comparative strength of ropes made of three different materials. They are taken from a table in Mr Kirkaldy's work giving the results of the testing of seventy specimens of ropes.
| Circumference. | Breaking or Ultimate Stress. | |
|---|---|---|
| Common hemp, untarred..... | 2.56 in. | 4840 lb. |
| Hemp, tarred..... | 2.69 " | 4263 " |
| Manilla..... | 2.62 " | 7041 " |
| Cotton..... | 2.48 " | 3089 " |
In these instances the strength of the untarred hemp rope is below and that of the manilla rope is above the average. Mr Kirkaldy states that the calculated 'stress per fathom-weight,' by a method akin to calculating the tensile strength per square inch of metals from actual tests, gives a more useful basis to compare the value of ropes than the breaking stress does. This stress per fathom-weight in each of the above examples was : hemp, untarred, 3457 lb. ; hemp, tarred, 2631 lb. ; manilla, 6905 lb. ; cotton, 2860 lb.
Ropewalk Spinning.—Notwithstanding the successful application of machinery to the manufacture of ropes, the old process of ropewalk spinning is still practised on a considerable scale. The successive stages in the process are (1) heckling the fibre ; (2) spinning the yarn ; (3) tarring in ' hauls ' consisting of about 300 yarns, laid close and parallel in lengths of say 100 yards ; (4) winding the yarn on bobbins and mounting these on bobbin-frames ; (5) forming the strands ; and (6) laying the strands into a rope. The heckling of hemp is done in the same way as the heckling of flax (see LINEN), the object being to remove the tow or short fibres and to place the long fibres called 'line' parallel to each other. Spinning in this process is done by hand, but either an iron whirl-hook, forming the spindle of a small pulley, or, what is of more recent introduction, a similar hook with a small cone upon it, is kept in motion for the spinner. Several of the former are mounted on a circular frame, and driven by hand or power ; but a set of the latter, which automatically fall out of gear by weights when the spinning is interrupted, are arranged in a straight line, because they are driven by the friction of larger cones fixed on a shaft, and there may be twelve of either kind of hooks forming a set. The spinner wraps a quantity of the heckled to the strand. In this way three or more strands are formed at the same time by the machine. These are then laid into a three-stranded rope or 'hawser' by attaching them at one end to the centre hook of the machine, while the other extremities of the strands are attached to three hooks. At that end where they are hung together on one hook the three strands are kept equidistant by placing them into the three longitudinal grooves of a conical piece of wood, called a 'top.' The twisting of the strands is effected by the rotation of the hook, from which the top recedes as the rope is formed. As the twist of the laying is in the opposite direction to the twist of the strands, the single hook requires to be turned in a contrary direction to the other three. A hawser has either three or four strands, the latter being said to be 'shroud hawser laid.' It requires a core-piece, and is much used for trawling. A Cable (q.v.) is a thick rope with usually nine, sometimes twelve strands.
Rope-making by Machinery.—By this is understood the making of ropes by machines which do their work without the necessity of having a rope-walk. The heckling, the drawing, and the spinning frames for preparing rope yarns are the same in principle as the corresponding machines used for spinning linen yarn for weaving purposes. There is, however, some difference in detail, owing to the greater average weight of rope yarn, so that, for example, there are comparatively few bobbins on the spinning-frame proper. Separate machines are perhaps more generally used for making strands and for laying these into ropes than compound machines which perform both operations, especially for very thick ropes. All these are now made of different designs, and with a good deal of variety in their details.
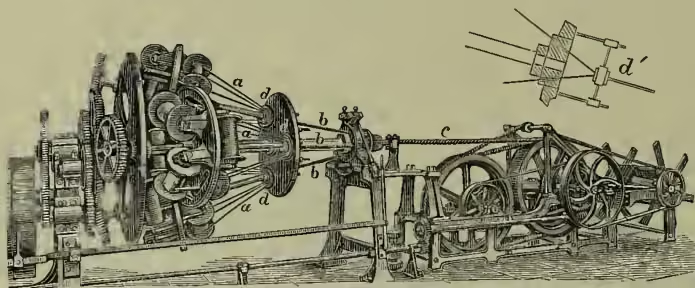
The annexed figure of Glover & Guitinane's patent compound rope machine will give a fair idea of one of the newest forms of this kind of apparatus. It is constructed to make three-strand ropes up to inches in diameter, there being in the one here figured six bobbins for each strand. It is furnished with change wheels, to enable either hard or soft laid ropes to be made. From a fuller description in The Engineer (7th March 1890) we extract the following : 'The yarns being wound round on the bobbins in suitable numbers, hemp round his waist, and attaches some of the fibres to one of the hooks, which by its revolving motion twists them as he continues to pull out and regulate the supply of fibres with one hand, and press them into proper form with the fingers of the other. He carries in his right hand a piece of woollen cloth, with which he grasps the fibres, and walks backwards, while he spins, to the farther end of the long covered walk.
The hauls of yarn are tarred by passing them through Archangel tar, heated to , a nipping apparatus being used to regulate the quantity taken up by the fibre. The next step is to wind the yarn on bobbins, which are then placed on a vertical frame. In order to form a strand of say ten yarns, one from each of ten bobbins is drawn through as many holes in a metal disc or register-plate, and immediately afterwards brought together and formed into a compact bundle of yarns by passing them through a stranding tube. On emerging from this the strand is fastened to the rotating hook of a forming machine on a travelling carriage, which, by means of proper gearing, is drawn along the walk, giving at the same time the proper twist according to the size of the rope to be made, they are from each bobbin threaded through a head-runner (register-plate) of six holes, and gathered at a die, at which they are closed into strands, there being a separate die for each of the three strands. The strands being formed, they are then threaded through a main head-runner of the holes, and immediately closed at the main closing die into finished rope. The rope is drawn through the die by means of strong hauling-off drums, and ultimately wound on a storage creel.—Wire-ropes (q.v.) and textile fibre ropes are now largely employed for driving machinery instead of belting.