Stem, the ascending axis of a plant, which usually bears leaves and flowers, and maintains communication between the roots and the leaves. In the Thallophytes—seaweeds, liverworts, &c.—no stem is differentiated; it begins among the mosses, grows stronger in the ferns, horsetails, and club-mosses, and attains its highest development in such trees as pine, palm, and oak. In these trees and in all other Phanerogams the stem results from the growth of the embryonic plumule.
Stems vary greatly in general habit; they may be upright and unbranched as in palms, or upright and much branched as in the oak; they may be scramblers as in the bramble, or twiners as in the hop, or climbers as in the ivy and Virginian creeper; they may be prostrate as in the strawberry, or underground root-like 'rhizomes' as in the Iris. Their usual function of lifting the leaves and flowers off the ground into the fresh air and sunlight may thus be lost, or subordinated to some other function, such as storage of nutritive material, as in the 'corns' of the crocus and 'tubers' of the potato, or storage of water, as in some succulents. When the rind is green it may assimilate as leaves do, and this is very important in such stems as those of cactuses, whose leaves are reduced to spines. It may also happen that branches of the stem are modified into flattened, leaf-like phyllodes as in Ruscus, into thorns as in the hawthorn, into tendrils as in the vine.
The stem is in many ways markedly contrasted with the descending axis or root, but few of the distinctions are rigid. Thus, most stems have some chlorophyll, which is absent from roots except perhaps in the case of one or two of aerial habit. Stems usually bear leaves, which roots never do. The tip of a stem is almost invariably a naked-growing point, while that of a root is usually protected by a root-cap. The branches of a stem arise as superficial outgrowths (exogenous buds), while rootlets arise endogenously from rudiments which develop in an internal layer known as the pericycle. The stem has a persistent tendency to grow upwards, while the root seeks the centre of the earth, and in regard to other forces than that of gravity, stem and root usually behave in opposite ways (see PLANTS, MOVEMENTS OF).
In describing aerial stems we distinguish the nodes from which the leaves arise from the intervening internodes, the buds which appear in the axils of leaves from that which forms the apex, or from those which appear arbitrarily or adventitiously, the leaf-bearing branches from the flower-stalks, and so on. The branching of the stem is usually lateral, but there are divergent forms, such as false dichotomy in the mistletoe, false axis in the vine, or true dichotomy in some Lycopods (see BRANCH).
In order to present a clear picture of the internal structure of a stem, it will be convenient to restrict our attention in the first place to the young twigs of some Dicotyledonous tree, such as the oak.
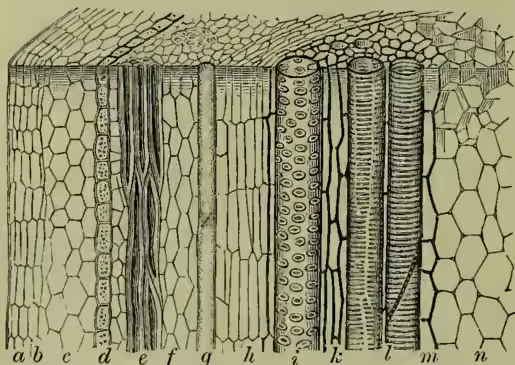
Every one knows that it is easy to peel off the 'bark' and to leave the white wood bare. This is possible because the region known as the bark is separated from the internal wood by a cylinder of delicate, readily ruptured, actively dividing cells—the 'cambium.' When a ring of bark is cut off a tree, the leaves do not wither; therefore we conclude that the water which ascends from the roots passes up by the wood. But if in the 'ringing' the young wood be also cut, the leaves wither rapidly; therefore it is by the young wood that the water ascends—a conclusion corroborated by the fact that a tree may flourish well although its heartwood has rotted away. But the wood includes many different kinds of elements—long vessels, tracheide cells, wood-parenchyma, and wood-fibres. It is certain that the younger tracheides and vessels are the paths for the ascent of the water. Again, if we tie a string very tightly around a stem so as to compress the bark, the stem sometimes swells just above the stricture; and if there be a fruit growing from the stem on that region, it will increase greatly in weight. This suggests that the nutritive materials elaborated in the leaves pass down outside the wood. But the tissue outside the wood and the cambium cylinder includes many different kinds of elements—an external epidermis, perhaps some cork, some softer rind or cortex, a set of hard bast-fibres, and, most internally, what is called soft-bast, including long 'sieve-tubes' and also 'cambiform-cells.' It is certain that this soft-bast is very important in the downward passage of elaborated sap.
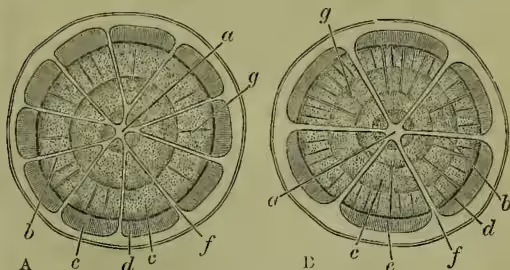
If we examine—by means of thin sections—the delicate growing point of the stem, we find that it consists of an external epidermis and of an almost homogeneous 'fundamental tissue.' As we pass in our examination from the tip downwards—that is, to slightly older parts—we notice that within the fundamental tissue there gradually appear certain firm strands. These differentiations of the fundamental tissue are known as fibrovascular bundles. Thus in the stem we distinguish the epidermis, the fibro-vascular bundles, and what remains of the fundamental tissue, in the form of central pith and superficial cortex, and radiating plates or medullary rays extending between these. In Dicotyledons the fibro-vascular bundles are radially arranged, and each consists of an internal wood or xylem part, an external bast or phloem part, and between these a persistent young layer of cambium, which, by the division of its cells, adds internally to the wood, externally to the bast, and enables the stem to grow in girth. These fibro-vascular bundles are continued out into the leaves, or, to state the fact in another way, each leaf contributes to the stem a 'leaf-trace' of fibro-vascular bundles which extend vertically down the stem, and eventually unite in a connected system. In the lower or older parts of the twig or stem there are necessarily very many fibro-vascular bundles, and these are intimately fused with one another. Thus we have from without inwards, beneath the cortex, a sheath of bast, a cylinder of cambium, and the central wood. The fundamental tissue remains undifferentiated in the cambium, and slightly modified in the central pith (if that be present), in the radial medullary plates which traverse the wood and bast, and in the superficial cortex.
The fibro-vascular bundles in the stem of Dicotyledons have a radial arrangement; in their vertical course they run parallel to one another, and are united into a connected system. Almost always they are 'collateral'—i.e. the bast lies in the same radius as the wood—and they are 'indefinite,' for the cambium persists. In Monocotyledons the fibro-vascular bundles which enter from the leaves pursue a curved course—first inwards and then outwards again—in the stem. In cross sections they therefore appear scattered in the fundamental tissue; they are also collateral but 'definite,' for the cambium is at an early stage used up. This last peculiarity explains the generally uniform thickness of a Monocotyledonous stem and the fact that the bark is not usually separable. In Ferns the bast surrounds the wood in each strand, forming 'concentric' bundles. So too there are structural peculiarities in the stems of Lycopods and Horsetails, while those of Mosses are, as we would expect, almost wholly cellular without differentiated bundles.
It is not easy to form an accurate conception of the internal structure of stems, but we must bear in mind the following physiological considerations. (1) As the stem is a supporting axis, often bearing heavy weights and resisting great strains, there is usually much skeletal tissue—e.g. the bast-fibres, the wood-fibres, and the hard wood which forms the heart of most trees. Schwendener and others have shown that the fibro-vascular bundles are always disposed to mechanical advantage. (2) As the outside of the stem is exposed to the weather and is often subjected to considerable strains, there is usually more or less protective tissue—e.g. the tough skin and a layer of cork. (3) As the stem mediates physiologically between the roots and the leaves, it contains conducting tissue—(a) the young tracheides and wood-vessels by which water and dissolved salts ascend; and (b) the sieve-tubes, cambiform cells, and conducting parenchyma by which elaborated materials descend. In the pith, medullary rays, &c. these materials are often stored.
The most practically important stems are those of timber-trees, but their uses are manifold. Thus, the bark of oaks (containing tannin) is used in tanning, and several kinds of bark are used in medicine; the bast-fibres of jute, hemp, &c. are used in making textile fabrics, ropes, and the like; the farinaceous pith of one of the palms is well known as sago, and many other kinds of pith are nutritious; the juices of some stems furnish india-rubber, oil, wine, &c.; the potato is the most important of the edible stem-tubers.
See BUD, BRANCH, LEAF, ROOT, VEGETABLE PHYSIOLOGY; H. Marshall Ward, The Oak ('Modern Science' series, Lond. 1892), for best English account of internal structure of stem, and Timber and its Diseases ('Nature' series, Lond. 1889), for theories of ascent of sap; Kerner's Pflanzenleben (vol. i., Leip. and Wien, 1889), for adaptations of stems; also text-books of Sachs, Van Tieghem, Asa Gray, Wiesner, and others.