Bird. Birds are in some ways the highest of the vertebrate animals. They represent the climax of that passage from water to land which the back-boned series illustrates. Their skeleton is more modified from the general type than that of mammals; their arrangements for locomotion, breathing, and nutrition are certainly not less perfect; their body temperature, higher than that of any other animals, is an index to the intense activity of their general life; their habitual and adaptive intelligence is familiarly great, while in range of emotion and sense impressions they must be allowed the palm. It is, in fact, only when we emphasise the development of the nervous system and the closeness of connection between mother and offspring, that the mammals are seen to have a right to their pre-eminence over birds. Birds and mammals represent two divergent lines of progress, and stand in no close connection, but the affinities between birds and reptiles are sufficiently marked to warrant their being included in a common class (Sauropsida), in contrast to the amphibians and fishes (Ichthyopsida) on the one hand, and Mammalia on the other. Among the numerous points of difference which separate birds from their nearest relations the reptiles, and from mammals, the following may be noticed:
| Reptiles. | Birds. | Mammals. | |
|---|---|---|---|
| Covering..... | Scales or scutes. | Feathers. | Hair. |
| Number of fingers..... | Always more than three. | At most three. | Five or fewer. |
| " skull condyles.. | One. | One. | Two. |
| " aortic arches.. | At least two. | One, right. | One, left. |
| Diaphragm..... | Only incipient. | Only incipient. | Complete. |
| Blood ..... | Cold. | Hottest. | Warm. |
| Position of optic lobes .... | On top of brain. | At sides of brain. | Covered up. |
| Parturition..... | Ovi- or viviparous. | Oviparous. | Except two, viviparous. |
But those contrasts are only a few of the less technical selected from Professor Huxley's masterly comparison of the three classes. To appreciate the full extent of the resemblances and differences between birds and reptiles, and the contrast between both and mammals, the reader must consult Huxley's Anatomy of Vertebrate Animals.
History of Study of Birds.—As birds are even to a careless eye most markedly separated from all other animals, there has rarely been any difficulty about the limits of the class. Aristotle (300 B.C.) defined them as feathered flying bipeds which lay eggs, and catalogued a goodly list, divided into three great orders, with the ostrich and a few others in separate subdivisions. He did not include bats in his list. For eighteen centuries no real progress was made with the study of birds. They were not of course lost sight of, but were abundantly discussed, now from a semi-religious, now from a practical point of view, yet it was not till the outburst of naturalist enthusiasm in the 16th century that they became again objects of scientific study. It is indeed curious, after a lapse of 1800 years, to find William Turner describing the birds enumerated in the catalogues of Aristotle and Pliny. Of the school known as Encyclopædists, Bêlon (1555) devoted himself especially to the study of birds, and made the great step of comparing the skeletons of bird and of man bone for bone. In the subsequent classificatory enthusiasm inaugurated by John Ray, and perfected by Linnæus, birds came in for a full share of attention, but the numerous naturalists who, towards the end of the 18th century, tried to put the birds in order, have left in their discrepant systems a record of the difficulty of a task which is still acknowledged to be bewildering. They fixed their attention mainly either on superficial characters, such as bills and feathering, or on general facts of habit, such as running and swimming. In other directions, however, progress was made: John Hunter (1728-93) and Vieq d'Azyr (1748-94) made important investigations on the skeleton and musculature of birds, while Caspar Friedrich Wolff, turning in 1764 to the problem of chick-development, the study of which had begun in Greece more than nineteen centuries before, laid the foundations of modern embryology. Then followed the Cuvierian school of anatomists, and to the works of the master himself, and of his numerous disciples, most of our present knowledge of bird anatomy is due. Numerous attempts at classification were again made, but even yet no one detailed system is generally adopted. General ornithology has been greatly enriched by the monographic, and in some cases stupendous labours of enthusiasts like Naumann, Thienemann, Audubon, Gray, Wilson, Gould, Sclater, &c. In recent years we are mainly indebted to Owen and Marsh for knowledge of fossil forms; to W. K. Parker for embryology; to Sclater, Wallace, Newton, and others for distribution; and for a scientific classification to Professor Huxley.
Structure.—(a) With the general form of birds every one is familiar; the frequent length of the neck, the inconspicuousness of the fore-limbs when at rest, the length of the hind-limbs, the reduction of the tail, the boat-like form of the main body, the smooth gracefulness of the contour curves are at once evident. (b) As to the appendages, the modification of the fore-limbs for flight in the great majority of birds is the most marked characteristic. While the flying reptile, Pterodactyl, had a membrane stretched on the much elongated little finger, and the bats have their skin spread out between their elongated four fingers, the bird has never more than three fingers, and these much reduced. The stroke, as every one knows, is effected not by a stretched skin, but by the strong stiff feathers. The most marked general feature in the legs is the elongation and fusion of the bones just below the ankle (metatarsals).
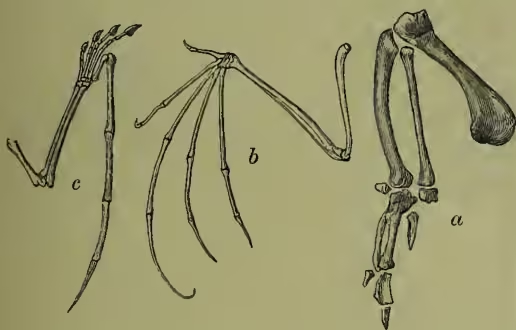
The toes are never more than four, except in abnormalities. The spur of the cock is thought by some to represent a toe. (c) The integument differs markedly from that of other animals in always consists of seven vertebrae, that of birds includes a larger and variable number, amounting to over a score in the swan. Unlike the very movable neck, the dorsal region tends to become more or less rigid, especially behind, where a large number of vertebrae enter into immovable connection with the hip-girdle. Behind this there are a few caudal vertebrae which are again mobile, and end in a compound ploughshare-bone which bears the great steering feathers of the tail. The most anterior ribs are fused to the vertebrae; then follow a few (two in the fowl) which are free, but do not reach being clad with feathers. These are skin structures generally comparable to scales or hairs. Their development will be discussed separately (see FEATHER), but it may be noted that in consequence of certain conditions of skin growth, numerous elevations are formed both of the outer and inner layers. These growing outwards, and at the same time fixed below in little pits, become to a large extent horny, and are split up in various ways to form feathers. The outermost sheath becomes a horny case which falls off as the feather grows; the feather proper is due to a horny modification of the inner stratum of the outer skin or epidermis; the inner skin (dermis) forms a nutritive base which, with its blood-vessels, keeps the young feather alive, but becoming subsequently reduced, leaves the perfect structure virtually dead. They do not usually occur all over the body, but in definite feather-tracts, as can be readily seen by plucking small or young birds. Some birds, however, like the penguin are really feathered all over. Three distinct kinds of feathers are at once distinguishable—(a) the small hair-like downy rudimentary filo-plumes; (b) the numerous smaller contour or covering plumes; and (c) the large strong quill-feathers or pennæ on wings and tail. The ordinary feather consists of a quill at the base of a shaft up the centre, and of the vane borne on the sides of the shaft. The vane consists of parallel barbs, which are linked together by small barbules. The base of the quill bears a miniature feather or after-shaft, which in a few cases becomes as large as the main feather. On the bare legs of many birds the feathers are replaced by horny scales, and the horny structures forming the beak and terminating the toes are very familiar. Skin glands, so abundant in mammals and elsewhere, are here absent; but at the very end of the tail there is a two-lobed oily preen-gland, the secretion of which is used by the bird for anointing and waterproofing its feathers. Curious skin structures, such as combs and wattles, are often developed.
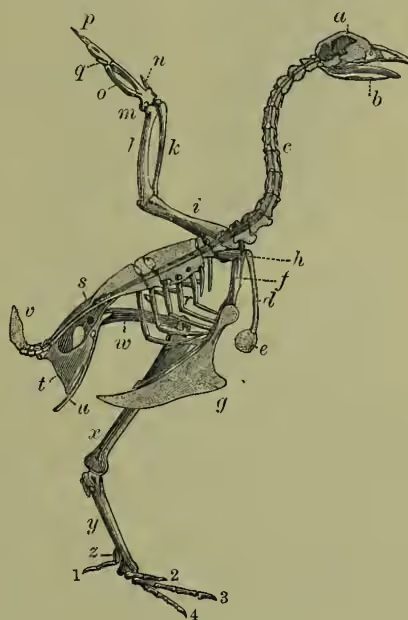
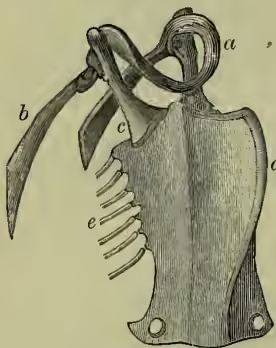
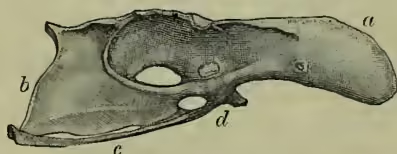
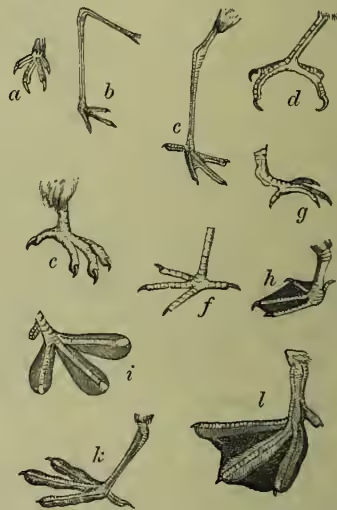
Skeleton.—The most important peculiarities of the bird skeleton are the modifications of fore-limbs and girdle in association with flight, the abundant air-cavities (only absent apparently in the kiwi or apteryx), which make the bones so light and spongy, and the tendency the bones have to fuse together. (a) The backbone is very different from that of other animals. The several backbone bodies (vertebræ) have a very complex shape, and are not to be mistaken for those of any other vertebrate. While the neck of mammals almost the breastbone; the next set meet, at an angle, upward processes from the breastbone, with which they are thus indirectly connected. The last ribs are again free. Almost all the ribs bear backward directed (uncinate) processes which overlap the rib behind. These also occur in crocodiles. (b) The skull of course corresponds in general structure to that of other vertebrates, but is distinguished by the early fusion of the component bones, so that the contours are no longer visible, by the prolongation of the front bones to bear the beak, by the absence of teeth, by the large sockets for the eyes separated from one another only by a thin (sometimes perforated) partition, and by the marked lightness of the bones. There is only one ball (condyle) which turns on the cup of the body of the first (atlas) vertebra, the bone (quadrato) on which the lower jaw works is movable, the portion of the skull lying in front of the eye-sockets is slightly movable on the portion protecting the brain, the tongue-bone (hyoid) is very well developed. It is on the relations of certain bones of the skull that Huxley bases his anatomical classification. (c) The limbs.—The shoulder-girdle consists (1) of a long bone running backwards (the shoulder-blade or scapula), (2) of a shorter bone running from the shoulder down to the breastbone (the coracoid, only a process of the scapula in mammals), and (3) of a slender collar-bone (clavicle) which extends from the shoulder to meet its fellow in front of the breast, forming with the broad uniting plate the familiar 'merrythought.' The breastbone or sternum forms the base of attachment for the great muscles which work the wings, it is concave internally, and in most birds bears a prominent ridge or keel on the middle line in front. As this ridge serves for the fixing of the flight muscles, it is largest in the birds which use their wings most, and absent in those, such as the ostrich family, which use them either not at all, or very little. The flying keeled birds (Carinate) are in the immense majority; the running birds, with flat raft-like sternum (Ratitæ), are few. Even among birds of flight, comparative disuse of wings may be associated with a slight development of the keel. In the New Zealand parrot (Strigops), and a few more, the keel is very rudimentary. The arm consists as usual of a strong upper bone or humerus, working in the socket formed by scapula and coracoid, of two bones forming the forearm, the radius and the stronger ulna, of a single pair of wrist-bones, and of three fingers. The three bones corresponding to the palm of the hand (the metacarpals) are fused together, the thumb has two, the first finger three, the second finger one or two joints. The terminal joints of the thumb and first finger are sometimes clawed. In the kiwi and cassowary there is only one complete finger, apparently the second. The feathers attached to the hand are usually called primaries, and those borne on the ulna, secondaries. The hip-girdle is very long, and its most important part consists of two dorsal bones (ilia) which are fused to a large number of posterior backbone bodies. A second portion (the ischium) extends backwards on either side, nearly parallel to the hind part of the ilium with which it is firmly fused. The third part (or pubis) also runs backwards from the socket of the thigh, and is a slender bone nearly parallel to the former. Only in the ostrich (Struthio) do the two pubes really unite; but in other birds they are bound together by fibrous tissue. In the American ostrich (Rhea) the two ischia are united ventrally. The socket formed by the union of the three portions of the hip-girdle is incompletely bony (perforate), as is also the case in one of the lowest mammals, the porcupine ant-eater, or Echidna. The thigh (femur) is a short stout bone. It is followed by the two bones of the lower leg, one of which (the fibula) is always imperfect and fused to the strong tibia. The knee is generally protected by a knee-pan or patella. The ankle is peculiar in this, that the upper portion unites in the adult bird with the lower end of the tibia, which is therefore known as the tibio-tarsus, while the lower portion unites with three of the bones corresponding to the sole of the foot (metatarsals), which are therefore known as the tarsometatarsals. In the adult there are thus no distinct ankle bones or tarsals, these having become fused to the tibia above, and to three of the metatarsals below. Birds have only four toes, and the metatarsals of the second, third, and fourth are fused together. The metatarsal of the great toe remains separate, but in many birds this digit is wholly undeveloped. The ostrich has only two toes. In most cases the number of joints in the toes increases by one, from two in the first to five in the fourth toe. The toes vary considerably in direction according to the way they are used. A bony spur ensheathed with horn is sometimes developed (as in the cock) in connection with the metatarsus.
Muscular System.—The most important muscles of the bird are those which work the wings. These lie on the breastbone, which forms their base of attachment. The great pectoral muscle which forms the greater part of the breast depresses the wing; a second smaller and deeper muscle, covered by the former, is continued into a long tendon which passes through a hole at the shoulder-joint, is inserted on the back of the upper arm, and raises the wing; a third much smaller breast muscle also assists in elevating the wing. It is well known that birds may remain firmly perched when asleep. This is effected by an automatic muscular arrangement. The tendon of a muscle which bends the toes passes up to the hip over the knee-joint, so that when the knee is bent in perching, the tendon is stretched, and the toes are kept firmly fastened. There is a strong development of muscles in association with the gizzard or stomach-mill.
a, fore brain; b, cerebellum; c, optic lobes; d, medulla oblongata; e, spinal cord; f, olfactory lobe; h, pituitary body; i, pineal gland.

Nervous System.—The brain of birds is markedly more developed than that of reptiles, the parts no longer lie almost on one plane, and the organ occupies the whole of the spacious skull-case. The convolutions characteristic of the fore brain of mammals are not yet developed. The 'tween brain (optic thalami) is quite hidden, the mid brain (optic lobes) is thrust to either side, the hind brain (cerebellum) exhibits cross grooves. There are twelve pairs of nerves given off as usual from the brain. The spinal cord exhibits in the hip region a conspicuous enlargement due to the divergence of its two sides. The sense-organs.—In association with their manifold experience, birds have well developed sense-organs. This is especially true of the eyes. They are large and powerful, and can be rapidly accommodated to different distances. They differ only in minor points from those of any vertebrate. The form is far from spherical, cal, the front portion being protruded, and to a certain extent constricted off from the ball behind. A special ring of small bony plates protects the base of this anterior portion. A comb-like nutritive fold (pecten), absent only in the kiwi, projects from behind into the posterior chamber of the eye. A third transparent eyelid or nictitating membrane (not restricted to birds) is conspicuously developed, and can be drawn like a screen across the eye. It lies hidden in the anterior corner. Tear and other glands are present. The ear differs markedly from that of mammals in the absence of any external portion. In the birds of prey, however, a membranous flap is present. The external opening is usually surrounded by a circle of fine hair-like feathers. As regards ear bones, only one (the columella) is present, and in this particular also the bird ear differs from that of mammals. The snail-like coil of the ear-cavity (the cochlea) is also much simpler. The (Eustachian) tubes which lead from the ear-aperture down to the mouth, unite before they open into the latter. The nostrils are usually situated at the base of the beak, but may occur further forward, and in the kiwi are situated at the very tip. In some cases they are guarded by bristles, in other cases prolonged into a tube, but are usually simple apertures. The nasal cavities may communicate with one another, and contain two or more gristly or bony scrolls. A peculiar nasal gland is present. The sense of smell is undoubtedly less developed than those of sight and hearing. Taste is also but slightly developed, and is discharged by the nerve terminations on the tongue, and by a soft patch of skin (ecere) usually situated on each side of the base of the beak. In rare cases (e.g. in mud-grubbing birds) a much larger area of the Bill (q.v.) may remain soft and sensitive.
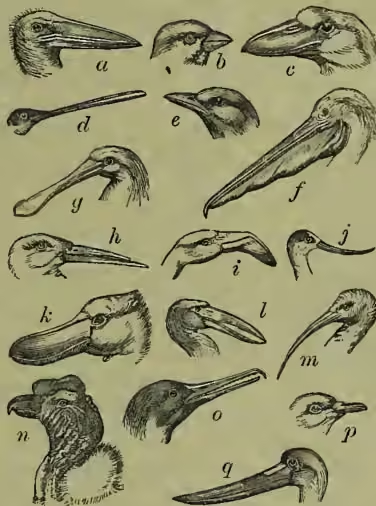
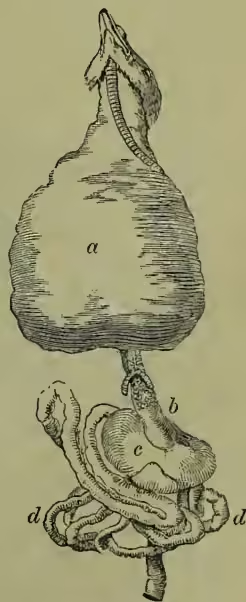
Alimentary System.—The character of the food-canal differs widely in different birds, varying with the nature of the food. Even in one bird the difference of food at different seasons, such as the change from fish to grain in the gull, may be associated with a marked change in the nature of the stomach. The beak may be helped by the tongue in taking up food, but in the absence of teeth no real mouth mastication takes place. The tongue varies widely in form, being not unfrequently notched and rough, and varies in size from the rudiment found in the pelican to the long protrusible organ of the humming-bird. Salivary glands open into the mouth-cavity, which in some cases (e.g. pelican) is enormously distended, and can hold quite a store of food. The gullet is soft and elastic, in obvious association with the downward passage of unmasticated food. In many birds it dilates into a spacious thin-walled sack, known as the crop, in which large quantities of food can be stowed away. It is from the crop that pigeons obtain the soft milky stuff with which they feed their young. In most birds the succeeding portion of the canal is divided into two (and sometimes three) distinct parts—the glandular and the muscular stomach. In the first (the proventriculus) the food begins to be digested, in the second (the gizzard) it is subjected to grinding. The former is characterised by thick, soft glandular walls, and near it the blood-gland, known as the spleen, is situated. Most people are familiar with the large hard gizzard of fowl or pigeon, with its very thick muscular walls lined internally by firm horny skin. The cavity usually includes small stones, and by the approach of the two sides the food is very effectively ground. The changes dependent upon food, above referred to, are expressed in the varying preponderance of glandular and muscular regions. The exit from the stomach is turned upwards (as in crocodiles), so as to lie very near the entrance. Of the long small intestine and the short large one, of the blind processes given off at the commencement of the latter, of the associated liver (very often without a gall-bladder) and pancreas, there is little of much interest to notice. The gut ends in a common chamber or cloaca, which also receives urinary and genital products. The back wall of this common chamber is connected with a peculiar sac with glandular walls, especially prominent in young animals, but of dubious import.
Vascular System.—The heart of birds shows a marked advance over that of reptiles, in the complete separation of arterial and venous blood. Like that of mammals, it is perfectly four-chambered. The blood-vessel arrangements also resemble those of mammals in the presence of a single main branch (aortic arch) conveying pure blood from the heart to the body; but this runs to the right instead of to the left. The most remarkable physiological fact about the blood is its high temperature of 100° to 112° F., greater than that of mammals by 2° to 14°. 'The polished feathers are bad radiators, the air entangled among them forms a bad conductor, and thus the heat is kept in.' The usual 'blood-glands' are present. There is only an incipient partition (diaphragm) between the anterior and posterior portions of the body-cavity.
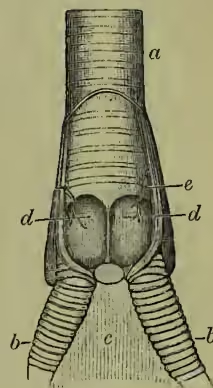
Respiratory System.—The two lungs do not lie freely in a closed space, as in mammals, but are fixed to the back of the bird. They are further peculiar in the way the supplying air-tube (bronchus) branches in a tree-like fashion without breaking up into finer and finer forking. The ends other, as John Hunter showed, with the air-spaces found in the spongy bones. The windpipe or trachea is often very long and coiled, and is strengthened by gristly or bony rings. It sometimes comes into very close association with the breastbone, running through part of it in the swan. At its upper end there is, as usual, a larynx; but this is not the vocal organ of birds. Further down, at the point where the windpipe divides into the two bronchial tubes, lies the seat of all the music, the sing-box or syrinx (see the paragraph on Song).
The Excretory System.—A pair of three-lobed kidneys embedded under the shelter of the hip-girdle perform the usual function of getting rid of nitrogenous waste. They are connected by a pair of ducts (ureters) with the cloaca. There is no urinary bladder. The urinary secretion is semi-solid, and rich in uric acid.
Reproductive System.—The male reproductive organs (or testes) are paired, and lie in front of the kidneys. The left is very often larger than the right. They are connected by a pair of ducts (vasa deferentia) with the common chamber, the wall of which occasionally bears a copulating organ. The female organs are peculiar in this, that in the adult the left side predominates over the right. In the embryo two ovaries are present; in abnormal and in a few exceptional cases (diurnal birds of prey) they may persist; usually, however, the right wholly degenerates. And similarly with the ducts, but the rudiment of a right oviduct is present. The end of the left oviduct is expanded to receive the ova as they burst from the ovary; the upper part of the duct is glandular, and equips the eggs with albumen; the lower is firmer, and contributes the limy shell; and the end opens into the cloaca whence the eggs are laid.
Nutrition.—The food of birds varies greatly, not only in different birds, but also to some extent at different seasons. Some are vegetarians, feeding on the green parts of plants, and in these the intestine is as usual long. Some confine themselves to grain, and these have large crops and powerful gizzards, while others combine cereals with insects (usually then without crop), or with the green parts of plants. A few sip honey, and a large number feeding on fruits probably play an important part in the dissemination of seeds. In insectivorous and fruit-eating birds the crop is usually absent, and the stomach only slightly muscular. Others again are notoriously carnivorous, or feed upon fish and molluscs, and in these cases the glandular predominates over the muscular portion of the stomach. The kea (Nestor) of New Zealand, originally a fruit-eating bird, has recently developed an alarming fondness for the blood and fat of living sheep. The interest of the nutrition of birds is twofold: on the one hand, in relation to the modifications of the alimentary system; on the other hand, in respect to the rôle of birds in the economy of nature. The experiments of John Hunter on the changes in the stomach of gulls consequent on a change of diet have been corroborated and extended. Such changes appear to occur normally in nature. Food is also said to have an influence on colour—e.g. in bullfinches and parrots.
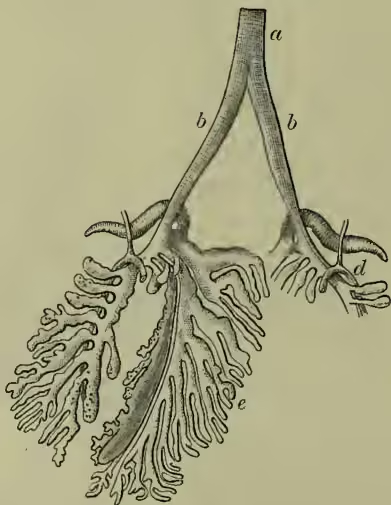
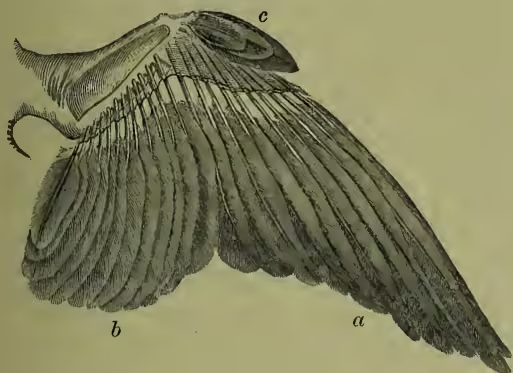
Locomotion.—Most birds use their wings in flight, the feather-covered arms being raised and depressed with great rapidity by means of the breast muscles already described. Every one who has watched birds is familiar with the marked differences in rapidity and mode of flight. It has been calculated that a common average of rapidity is about 40 to 60 feet per second, but records of the feats of carrier-pigeons, &c. certainly greatly surpass this. It seems probable that strong-winged birds, like eagles, can cover about 80 feet in a second. Buffon noted that they disappeared from sight in about three of the bronchial branches are in connection with air-sacs which lie round about the lungs. These greatly increase the quantity of air which may be taken in; they are of use in warming it, and doubtless aid, not only by greatly reducing the specific gravity, but also on the heated-air principle of Montgolfier's balloon, in buoying the bird in the air. These air-sacs are large but thin-walled bags, nine in number (see AIR-SACS). They are connected on the one hand with the bronchi, on the minutes. The importance of the air-sacs and spaces has been already noted. Strong birds, like the albatross and birds of prey, can not only fly very rapidly, but can sustain their exertions for long periods, while many other birds rarely take prolonged flights, except during migration. The ostrich uses its wings to help it along in its rapid race; some aquatic birds, like the steamer-duck, use them as paddles, auxiliary to its legs. On the ground, birds vary greatly in rate and manner of progression, the swift strides of the ostrich, the rapid run of the partridge, the hopping of the sparrow are well-known illustrations of different gaits. That many birds are expert divers and climbers is also a familiar fact. See FLIGHT.
Respiration.—The great activity of birds is associated with very efficient respiration. Expiration, or the expulsion of used air, is managed by the contraction of breast and abdominal muscles, which compress the inclosed cavities and force the air from the sacs and lungs. When these muscles are relaxed the cavities again elastically expand, and fresh air rushes in by the windpipe to lungs and air-sacs.
Song.—With few exceptions, birds have a vocal organ, and are able to produce more or less variable sounds. The organ is, however, wanting in the running birds, such as the ostrich, and in the American vultures. The sounds produced are almost as varied as the different kinds of birds, and an expert ornithologist has little difficulty in identifying a great number of forms by their distinctive noises. That some chirp and others scream, that chattering describes the language of many and croaking that of others, that some boom and others bark, that the crows caw and the laughing jackass laughs, that the mocking-bird imitates and the parrot becomes able to articulate, and above all that the lark trills and the nightingale truly sings, are well-known illustrations of the variety of bird language. The weird cry of the curlew or whaup, the melancholy voice of the seamew, the gabble of ducks, the crowing of the cock, the soft cooing of the dove, the hoarse voice of the corncrake, the ecstatic melody of the bobolink, the cheerful notes of the blackbird, the educated music of the canary, are again a random selection of instances from an almost infinite medley. It is among the so-called perchers, songsters, or Insectores, that we find song really developed, and that for the most part in the males, and in highest degree at breeding time. Though the notes are not musically pure, many bird songs have been expressed in musical notation, and every one is familiar with imitations in word form.
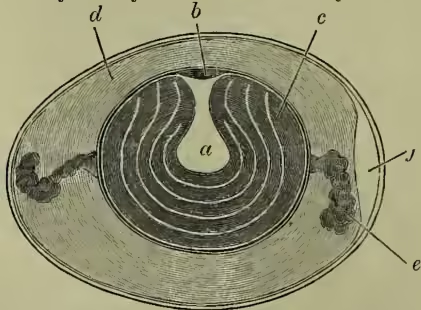
In mammals the voice is produced by the mechanism of vocal chords situated in the larynx at the top of the windpipe. In birds these vocal chords are never present, but their absence is compensated by the development of a sing-box (or syrinx) at the base of the windpipe. In some cases the sing-box is wholly produced from the base of the windpipe (or trachea)—e.g. in Thamnophilus and American Passerines; in a few cases the mechanism is confined to the beginning of the bronchial tubes (e.g. Steatornis); in the great majority of songsters the syrinx includes the base of the trachea and the tops of the two bronchi. The mechanism is a very variable one, and even in its simplest forms not to be understood by any verbal description unaccompanied by dissection of a few typical singers. To take a typical example of a lower larynx or syrinx of the last and commonest construction, we find (a) that the base of the trachea and the tops of the bronchi are altered by lateral compression or by expansion, by fusion of several of the strengthening rings, or by incompleteness of some of the same. (b) The end of the windpipe is divided by a bony ridge, rising up at the angle where the two bronchi unite, and forming the main stay of the resonating cavity known as the 'tympanum.' (c) From this bony ridge to the median surface of each bronchus stretches the 'internal tympaniform membrane.' (d) Between the last tracheal and the first bronchial ring, or between a pair of modified bronchial rings, stretches a second 'external tympaniform membrane.' (e) In singing-birds there is a third important inward projecting 'semi-lunar membrane.' (f) Besides these there are vocal membranes or folds of skin, projecting inwards in both bronchi. The tension of the membranes is varied by the action of special muscles; their vibration as the air passes over them causes sound; the note varies with their muscular state, with that of the tympanum, and with the column of air in the windpipe.
Singing is an unbidden expression of emotional energy. It is most marked at the high tide of sexual emotion during the breeding-season. It is best, sometimes solely, developed in the males, who use their powers to attract the females, and often vie with one another in so doing. In other cases the note is obviously used as language, expressing alarm and the like, for that some birds are able by voice to convey impressions to one another is indubitable. In so far as the song is an instrument and expression of sexual attraction, it falls to be included among those powers which have been strengthened and developed by sexual selection.
Moulting (q.v.).—After the strain of the reproductive period, or sometimes at the low ebb of mid-winter, the old feathers drop off, and birds undergo annual moult. The use of this in replacing breakage, and in furnishing a complete machinery for the flight of migration, is very evident; the cause is not yet sufficiently investigated. Moult obviously presents some analogies with skin-casting and hair-shedding in other animals, and must be associated with some deep-seated constitutional change, such as its connection with the end of the breeding-season suggests. Besides this annual growth of new feathers, many birds exhibit double, and some triple moulting. The ptarmigan, for instance, changes its suit three times in the year, moulting after breeding into gray, changing this for white as the winter sets in, and acquiring in spring a third and most attractive set of feathers. In association with sexual attraction many male birds seem to undergo a partial moult, as the result of which they acquire those special decorations which are the index of a reproductive climax.
Reproduction.—Birds usually pair in spring-time, but to this rule there are many exceptions. Fertilisation is internal, and all birds are oviparous. The number of eggs is often in inverse proportion to the size of the bird. Several, such as the apteryx, lay only one; the doves and the birds of prey lay two or three, but the majority of birds many more.
The Sexes.—The male birds are often markedly different from the female. In the Bird of Paradise (q.v.), for instance, the two sexes are at first alike, but with successive moults and increasing maturity, the male becomes in colour and plumage a most marked contrast to the comparatively sober female. The contrast between peacock and peahen is a yet more familiar illustration of a very widely observed fact. In rare cases the female is the more brilliant. The male has also a richer and more varied voice, and in many cases the singing is all on his side. Both with song and with dance, by strains of love and display of charms, by the wildest excitement of aerial evolutions, and by the most grotesque of striking attitudes, the males strive to captivate the females. For the details of this sexual attraction, and for the importance of sexual selection in evolving decorations and charms, the reader may be referred to Darwin's Descent of Man, where the subject is very fully discussed. Birds associate together to very different degrees; thus, eagles live separately, the two sexes only coming together at breeding time; the doves live in pairs; many birds are similarly monogamous, but live in society, except during the breeding-season, while fowls and some running birds are polygamous, one male ruling over a number of females. See SEX.
Eggs.—The size of the egg is very generally proportionate to that of the parent, but there are many striking exceptions. The very centre of the egg contains what is known as white yolk which rises as a plug to the surface. This white spot, usually lying uppermost when the egg is opened, is the area (blastoderm) where by the multiplication and arrangements of cells the embryo chick is mapped out. The 'yellow yolk' is traversed by concentric layers of white, which also forms a thin layer round the outside. As the egg passes down the upper portion of the oviduct it becomes coated with fluid white of egg or albumen, which is separated from the yolk by a (vitelline) membrane inclosing the latter. The fluid albumen is also disposed in layers, separated by fibrous networks, and almost every one is familiar with two thick sets of fibres (known as the 'tread') which extend from each end of the yolk into the surrounding albumen. In the lower portion of the oviduct (the uterus) the egg becomes, as we have already noted, inclosed in a limy shell. This is an organic structure and is two-layered. Its texture and microscopic characters differ in the various orders, and appear to be truly diagnostic. The shell is separated from the white of egg by a double membrane, the two layers of which become separate at the broader end, and leave a familiar air-space which increases with the age of the egg. The form of the finished egg varies from an approximately perfect sphere to more or less symmetrical ovals. In the uterus the shell usually becomes stained by complex secretions of pigment which in some respects resemble those of the blood and bile. As many as seven distinct pigments have been distinguished by Mr Sorby. It is necessary to distinguish the ground colour from the subsequent more or less circular markings, which become variously blotched and distorted by the movement of the egg in the uterus. The markings are not always constant, but vary with age and even in the same brood. The colouring is apparently to some extent protective against the sun rays, and varies also according to exposure. That the colour of the eggs often favours their being overlooked by enemies is a familiar fact. Though the older naturalists, before the collecting of eggs had been superseded by embryology, doubtless exaggerated the importance of their science of 'oology,' it is also certain that some important hints in classification have been afforded by the study of the characteristic colouring. The extraordinary enthusiasm exhibited by egg-collectors, from the time of Sir Thomas Browne's cabinet onwards, is a most remarkable side chapter in the history of science, which has been recapitulated in the lives of many of our famous naturalists, and indeed in the boyhood of almost every boy bred in the country. See EGG, CAGE-BIRDS, INCUBATION.
Development.—The development of the embryo bird will be discussed along with that of other backboned animals (see EMBRYOLOGY); but a few general facts (with special reference to the fowl) may be noticed here. The egg is fertilised in the upper part of the oviduct before the coating of albumen is added to the ovum proper. The usual division commences in the lower part of the oviduct, shortly before the shell has begun to be formed. Owing to the vast amount of nutritive, as opposed to formative material, the whole ovum does not divide, but only a special part of it, known as the germinal disc. It is also noteworthy that the division proceeds somewhat unsymmetrically. The subsequent changes by which the different germinal layers are established, and nervous system, backbone, gut, muscles, and the organs generally are formed, cannot be here discussed. The development of the chick excited interest even among the old naturalist philosophers of Greece, and we have not finished the inquiry to-day. It was by observation of the embryo chick that Wolff first showed distinctly (in 1759) that development was a matter of gradual growth, and not the unfolding of a miniature ready-made organism. It was in regard to the fowl that Pander (1817) first drew emphatic attention to the different germinal layers, the distinctions between which are of such importance in understanding the process of development. On the development of the chick also, Von Baer (1828-37) partly based his great generalisation as to progress from the simple to the complex, and showed that the passage of the embryo from one great stage to another corresponds to the successive steps which mark off fish from amphibian, amphibian from reptile, reptile from bird. This law we now express by saying that the individual development (ontogeny) is a recapitulation of the historic evolution of the race (phylogeny). Mr Parker eloquently compares watching the development of a bird to reading a palimpsest; under the more superficial characters are others (at an earlier stage) more primitive, beneath those others more primitive still, till the characters are seen to be distinctly reptilian, and so on backwards. The work thus begun by Wolff, Pander, and Von Baer has been vigorously continued; many of the greatest embryologists have contributed to the elucidation of bird development; in Britain Parker especially has, with marvellous patience and skill, followed the development of the bird skull. By the recent use of the contrivance known as the embryoscope, the development of the chick has been watched going on, and even experimented upon.
The chick is inclosed in an embryonic membrane known as the amnion, and has a large respiratory sac called the allantois. The nutritive yolk which is gradually absorbed is inclosed in a 'yolk sac.' When about to be hatched, the chick thrusts its beak through the egg-membranes, and breathes the air in the air-chamber. The lungs begin to work, and the allantois shrivels up. The Umbilicus (q.v.) becomes completely closed, and 'the chick, piercing the shell at the broad end with repeated blows of its beak, casts off the dried remnants of allantois, amnion, and serous membrane, and steps out into the world.'
The period of incubation, during which the parent, generally the female, but often also the male, sits in most cases constantly on the eggs, varies greatly, from ten days to eight weeks. The young birds may be at first naked, blind, and helpless, and are then fed by the parents till fledged (Altrices or Insessores)—e.g. Thrush, or they may be downy, active, and able at once to look after themselves (Præcoces or Autophagæ)—e.g. Chick. In some cases, however, they are hatched downy, but remain for a while helpless. See EMBRYOLOGY.
Nests.—It is important to notice that the higher development of birds, as compared with reptiles, is associated with the production of fewer offspring, but at the same time with the enormous increase of parental care and sacrifice. If the young are to be developed within the eggs, the latter must be kept at an approximately constant warmth. In almost all cases this is effected by brooding. The comfort of the parents during the hatching period is secured by the nest, which becomes further necessary for the protection of the fledglings. The nest-building instinct is not indeed the exclusive property of birds, but occurs also among insects and fishes, not to mention the numerous cases where homes rather than nurseries are constructed by animals. But it is among birds that the instinct reaches perfect development. The prolonged period of brooding, the frequent helplessness of the young, the very common arboreal habit, the not unfrequent enemies, have necessitated a most varied series of nest-building contrivances. The nest is built before the eggs are ready to be laid, and in most cases the female takes the prominent part in its construction. But both in the building and in the subsequent brooding the male may do his share, or in some cases much more. Each species usually has its own peculiar style and material of construction, though this may be adapted to varying conditions. The nests are usually solitary, more rarely grouped, and very exceptionally (as among cassowaries and ostriches) common property. Rooks, sea-fowl, herons, are familiar examples of breeding communities, while the sociable grosbeak, the republican swallows, and a few others, form even closer associations. The cuckoo and the cow-bird have managed by a sort of parasitism to shirk their task, and quite a number of birds lay their eggs in an exceptional manner in the nests of neighbours. The beak is the organ most used in construction, but the pressure of the body may round off the forming nest, and the feet may also be used. How comfortable a nest may be made inside every one knows; how adroitly hidden it may be by external decorations of moss and lichen is familiar to every nest-hunter. All who have opportunity should examine the cases of birds and their nests so beautifully arranged in the British (Natural History) Museum in London. The smaller birds usually build the more beautiful nests, and every variety occurs, from the comparatively careless hole in the sand made by the ostrich to the skilfully suspended and neatly fashioned nest of the tailor-bird.
Without entering into details of nest-construction, the following series given by Leunis may be noted. For details, such a work as Rev. J. G. Wood's Homes without Hands, and for theory, Wallace's essay on Nests in his Contributions to the Theory of Natural Selection, may be profitably consulted. It must at the outset be again noticed that habits vary considerably, as the very diverse nests built in different circumstances by falcon, eagle, heron, &c. well illustrate. Nests are shifted to suit food-supply, and vary in structure according to the available material. And again, since nest-building is obviously an acquired habit, which gradually rewarded the species in the greater success of both parent and offspring during breeding time, it is natural to find it dispensed with in many cases where the nature of the situation rendered no actual nest necessary, or where the birds for some other reason have never learned the habit. Some sea-birds, like the auk, simply lay on the rocky ledges of their haunts; some ground-birds simply deposit their eggs on the bare soil.
(1) Burrowed holes are made by sand-martins, bee-eaters, penguins, kingfishers, and many others. The prairie-owl, living in the burrows of the prairie-dog and of the armadillo, is a well-known example of peculiar habit, and in the first case of curious partnership. (2) Ground-nests, generally of the simplest character, with rough and scanty accumulation of nest material, are made by swans, ducks, geese, fowls, gulls, waterhens, corncrakes, &c. (3) Mud-nests, constructed from damp earth, are well illustrated by the house-swallows, blue-creeper, flamingo, &c. The common singing thrush is well known to make a firm nest of clay and cow-dung mixed with moss. (4) Carpenter-nests, formed with more or less preparation in the holes of trees, are used by woodpeckers and a few other arboreal birds. (5) Platform-nests, simply consisting of flat seats, are formed by the ring and turtle doves, by eagles, storks, and cranes. In some parts of the Continent the flat nests formed by the storks on the tops of buildings are familiar enough objects. (6) Basket-nests are such loosely interwoven constructions of grass, stems, twigs, &c., as are made by crows, missel-thrushes, and most singing-birds. The green weaving-birds (Plocicus pensilis) hang their loosely woven nests, with downward directed opening, on the Madagascan trees. The South African republican birds (Philectavus socius) form hundreds of hanging nests on the branches, under the shelter of a common thatch. (7) Woven nests are the more delicately constructed and really woven constructions of wool, hair, bark, grass, &c., which are made by such birds as the goldfinch, the Baltimore bird, and very many others. (8) Sewed nests, composed of leaves sewed together by the beak as needle, are well illustrated by various species of Icterus, and by the Indian tailor-bird (Orthotomus benetii). (9) Felt-work nests are woven from the wool of plants or animals, sometimes with other material in addition; the humming-birds and the bullfinch form beautiful nests of this fashion. (10) Cement-nests are bound together by a viscid and very adhesive secretion, which is mixed with saliva, and used to glue the materials of the nest together. The nests of the American swallow, the edible birds' nests of the Salangani, sought after as luxuries by Chinese and others, are of this cemented type. (11) Dome or moss nests are roofed in above, and have an entrance on the side. The common wren, the water-wagtail, and the tits build on this principle. The beautiful bottle-shaped nest of the titmouse is one of the best examples. (12) The parasitic habit is well known among cuckoos and cow-birds. The nest of another bird is utilised to the future loss of the rightful inmates, and with obvious economy of labour on the part of the intruders. Thus sparrows usurp the nests of swallows, and starlings those of woodpeckers. Pheasant and partridge eggs are sometimes found in the same nest, and the same has been observed in many cases—e.g. gull and eider-duck. When artificial nests are forthcoming, birds are glad to be relieved of the labour of construction, and different birds thus sometimes share a common box. The resorts of birds, when convenient nooks are available, are often extremely curious.
Migration.—It is a well-known fact that comparatively few birds (at anyrate, outside of the tropics) remain in the same place all the year round. They do not hibernate, but migrate on the approach of cold. Some we know as winter visitors returning north again in spring, most we know only in summer, for in autumn they fly to the warmer south; a third set we call 'Birds of Passage,' for these we only know somewhat incidentally as they pass through on their way elsewhere. Thus the swallow, the cuckoo, the nightingale, &c. come to Britain in summer and breed there, being winter residents farther south; the fieldfare, jack-snipe, bean-goose, redwing, &c. reach Britain in winter, being summer residents and breeders farther north; while the little sandpipers are familiar examples of the true birds of passage which we know only for a short time as they rest on our shores in their journey south in autumn, and north again in spring. These three classes are obviously only different cases of one fact of Migration (q.v.). As this will be discussed separately, only the general facts are here to be noticed. Almost all birds are in some degree migratory. Those which breed in the equatorial regions are the chief exceptions, and even they pass from hill to valley and back again. Forms, too, which seem to be constant residents of a non-tropical country are in many cases known to exhibit a partial or a very local migration. This is true, for instance, of the common wren and the red grouse in the north of Scotland. All birds breed in the colder regions of their migration. Changes in food-supply and temperature are the most important conditions impelling them to shift their habitats. The general trend of migration is always, as indicated, towards the equator in autumn, from the equator in spring; but the investigations of the British Migration Committee have clearly shown that the courses often come to be circular. The flight is the more universal in a country, the more marked the contrast between summer and winter. The annual migration from breeding areas too cold for winter residence and food-supply to warmer subsistence areas cannot be understood apart from the history of climates. When the European climate was more equable, it was virtually indifferent to the birds where they went. As it grew colder, the birds had to fly farther and farther south every few winters. Migration has become an inherited habit, for they set about it before the impelling conditions are directly present. According to Wallace, natural selection has played an important part in confirming this habit. Many facts about migration are still utterly obscure. The power birds have of flying straight and of returning to the same locality is very marvellous. It must be remembered that a continuous tradition is sustained; those who have made the journey before guide the others. Doubtless they have memory for great landmarks. They fly across the shallower parts of the Mediterranean, where a chain of islands in this submerged tract long remained to guide them. The smaller birds usually keep nearer the ground; but it must not be forgotten that the flight is usually mostly accomplished by night. Birds generally meet in concourses, and migrate in flocks. Only a few fly alone. Sometimes the old males remain, while the others 'flit.' The return northwards is more rapid, without young ones or weaklings. The males often return first.
Intelligence and Emotions.—As birds have a full active life, with considerable variety of function, in usually complex environment, since, as we have already noticed, their sense-organs and nervous systems are highly developed, considerable exhibition of intelligence is to be expected. They seem to have great vividness of sense impressions, to judge from their power of recalling old haunts and old friends. Birds often return to the same place season after season, and they have been known to recognise an owner after the lapse of years. Their quickness of ear and power of retention are evidenced by the power some possess of learning to repeat sounds, both words and tunes. Some have exhibited marked fondness for music, and the æsthetic tastes of the Bowerbird (q.v.) excite deserved admiration. Much more is known in regard to their marvellous hereditary, general, and largely automatic reasonable habits or 'instincts,' than in regard to their power of individually adapting their conduct to novel circumstances. Their beautiful and adroit contrivances of nest-building are very familiar instances of the former, but many instances of the latter have also been recorded. Mr Romanes has collected numerous illustrations of curiosity, imagination, forethought, and instruction by experience. He notes many interesting cases of their special devices in obtaining food, in avoiding capture, and in nest-building. His valuable treasury of sifted facts ought to be consulted on this subject.
As to feelings, it is hardly necessary to refer to their unexampled exhibition of sexual emotion in song and dance, parade and display, or to the marvellous parental love and sacrifice expressed in their nest-building labours, in their prolonged incubation, and in their care for and courage in defending their brood. Subtler emotions of jealousy, both in connection with and altogether apart from sex, of affection for owners or associates, of sympathy for wounded or enfeebled fellows, are also not rarely exhibited. That a bird singing continuously for hours does not represent a rare height of emotion is not to be believed. It may be fairly said that the joyous song of the lark 'at heaven's gate' is an eloquent expression of emotion only surpassed perhaps by human music.
Distribution in Space.—It is a curious fact that though birds are able to fly far and wide, and are much less settled than most animals, it was in relation to them that the first great essay towards an understanding of distribution was made. In 1857 Mr Selater based, upon the facts of the distribution of perching birds (Passeres or Insectores), a division of the earth into six great regions, with characteristic fauna. These regions he termed Palearctic, Ethiopian, Indian, Australian, Nearctic, and Neotropical. For an account of the birds which especially predominate over these great regions, or others with somewhat altered limits, the reader must be referred to the works of Murray and Wallace on Geographical Distribution of Animals, and to the convenient summary of facts given by Heilprin (Inter. Science Series). Some genera of birds are very widely distributed, and almost universally represented; others, like the birds of paradise and the humming-birds, are peculiarly restricted. It hardly needs to be stated that birds have unusually great facility of dispersion; that they are more numerous in the warmer countries; that according to the nature of the region, different physiological types will preponderate—e.g. insect-eating birds in the tropics, and fish-eating swimmers in the northern seas.
Distribution in Time.—(1) In the Triassic strata of New England, various kinds of footprints occur which most naturalists now regard as reptilian, though others are inclined to see in some of them the earliest hints of birds. (2) The oldest known birds are found in the Upper Oolite, the well-known Archæopteryx (q.v.), from the Solenhofen lithographic slates, and Laopteryx priscus, from 'a nearly equivalent horizon in the Wyoming Territory' of the western United States. The long feathered tail, the unfused metacarpal bones, the toothed jaws, and other features of Archæopteryx, are exceedingly important pristine characters. (3) In the Middle and Upper Chalk, numerous fossil birds have been discovered, especially by Marsh. These are grouped together in a somewhat mixed division of toothed birds, or Odontornithes (q.v.). Of these the most remarkable are Hesperornis and Ichthyornis (q.v.), both with teeth. The latter had biconcave fish-like vertebrae, well-developed wings, and a keeled breastbone. The former had vertebrae much like those of modern birds, rudimentary wings, and, as one would expect, no keel on its breastbone. Its teeth were not implanted in distinct sockets as in Ichthyornis, but lay in a longitudinal groove of the jaw. Apatornis was another remarkable form, and Enaliornis has been found in Europe in the Upper Greensand of Cambridge. (4) In the Tertiary deposits birds become more numerous, and liker those now existing. Representatives of modern genera occur from the Eocene onwards. Many types, such as the great running birds, now with restricted distribution, are found in regions far from their present habitats. A giant goose-like form, Gastornis, apparently exceeding the ostrich in size, has been found in France and Britain. Heilprin notes how Leptosomus, type of a small family now restricted to Madagascar, is found in the Eocene deposits of France. In the Eocene also occur Megalornis, little smaller than an emu; Dasornis, uniting ostrich and moa; Odontopteryx toliapicus, 'with dental armature recalling the toothed birds' of the chalk. A crane-like genus, Aletornis, is frequently represented in American strata. Giant forms disappear, the running birds are driven into corners, teeth degenerate and wings develop, as the modern bird fauna is gradually approached. Those who wish to read the full story should consult especially the writings of Marsh, and in the first place convenient summaries, such as that of Heilprin.
Sub-fossil and Historically Exterminated Birds.—Within comparatively recent (semi-historic) times, the 'moas' of New Zealand (Dinornis and Mionornis), the Aepyornis maximus of Madagascar, the Australian Dromæornis (ancestor of the emu), and others have disappeared. Much more recently the dodo of Mauritius, the solitaire of Rodriguez, the crested parrot and the Aphanapteryx of Mauritius have been exterminated. Under separate articles some of these will be discussed. Thoughtlessness and cruelty are unfortunately tending to the extermination of some other forms.
Classification.—In 1880 Mr Selater estimated the number of extant bird species at 10,139, and Gray's estimate was somewhat higher. To these some 200 known fossils have to be added. The classification of these numerous forms is still a matter of the greatest difficulty. The older classifications were based on the habits, or on comparatively trivial characters; but as this physiological classification has been gradually improved, it has come to be not so inconsistent as formerly, with the anatomical classification suggested by Huxley. In his famous paper on the 'Classification of Birds,' Proceedings of the Zoological Society, 1867, Huxley grouped birds according to certain features of the skull, and though this may seem too narrow a basis, it must not be forgotten, as W. K. Parker remarks, 'that the structures of the skull and face govern the whole body, as it were; every other part of the organism corresponds to what is observable there.' But even when the great groups are marked off on this basis, there remains the huge difficulty of the genera and species. The difficulty of satisfactory diagnosis is due to the narrow range of modification. 'One bird is so like another,' say the non-ornithologists, and Professor Huxley confesses the same in saying that 'the structural modifications which the specific forms present are comparatively little importance; any two birds which can be selected differing from another far less than do the extreme types of the lizards.'
While there is general agreement in distinguishing three great divisions of (1) ancient birds, like Archæopteryx (Saururæ); (2) running keelless birds, like ostrich (Ratitæ); and (3) keeled flying birds (Carinatæ), the divergence of classification observed in grouping the last of these three divisions is very great. In many works, especially text-books, the old orders of swimmers (Natatores), waders (Grallatores), scratchers (Rasores), climbers (Scansores), perchers (Passeres or Insectores), and birds of prey (Raptores), are still followed. In other cases some of these are retained, and others split up or rearranged in smaller orders, and a dozen or more established in all. It seems therefore most useful simply to append the classification of Huxley, with examples showing their relation to more familiar divisions.
- I. Saururæ.—Metacarpals not fused. Tail longer than body. Often with teeth—e.g. Archæopteryx.
- II. Ratitæ.—None of preceding characters. Flat unkeeled breastbone—e.g. (1) the kiwi (Apteryx), with a big toe; (2) the 'moas,' cassowaries, and emus without a hallux; (3) the ostriches, differing from the preceding in having a long upper arm (humerus), and two (not one) clawed digits—the American ostriches or Rheas, with ischia (see hip-girdle) united, but pubes free, and the true ostriches or Struthios, with the ischia free, but the pubes united.
- III. Carinatæ.—Ordinary birds with keeled breastbone.
- 1. (a) Dromæognathæ.—Vomer broad behind, interpolated between pterygoids, palatines, and basisphenoid rostrum—the Tinamous.
- (b) (2-4) Vomer narrow behind, pterygoids and palatines articulating largely with basisphenoid rostrum.
- 2. Schizognathæ.—Maxillo-palatines free, vomer truncated in front. Plovers, gulls, penguins, cranes, fowls, sand-grouse, pigeons, hoazins, goatsuckers, humming-birds.
- 3. Agithognathæ.—Maxillo-palatines free, vomer truncated in front. Part of Gerauomorphæ, the Hemipods, swifts, Passerine birds.
- 4. Sauromorphæ.—Vomerine halves permanently distinct, maxillo-palatines arrested (except in woodpeckers, united more or less completely). Woodpeckers, birds of prey, parrots, cuckoos, kingfishers, trogons, goose-like birds, flamingos, storks, cormorants.
- 1. (a) Dromæognathæ.—Vomer broad behind, interpolated between pterygoids, palatines, and basisphenoid rostrum—the Tinamous.
While the meaning of the terms can only be appreciated with the skull in hand, the above classification is independently of value as showing what orders of birds are on anatomical grounds most nearly related.
Pedigree.—It is generally allowed that birds have risen out of reptiledom. Such an origin is suggested not only by the general fact that 'reptiles' in the wide sense form the nearest distinctly lower class, but by resemblances of structure revealed in anatomical research, and still more by the study of bird embryology. But the crowning evidence in favour of this origin is found in the primitive characters of the first known bird (Archæopteryx, q.v.), and by the series of forms connecting flying saurians and toothed birds. Birds are derived from the ancient reptiles or saurians known as Deinosaurs or Ornithoscelida. These lead up to the primitive birds or Saururæ, from which two branches diverge, represented by the keelless birds (Ratitæ) on the one hand, and by the keeled birds (Carinatæ) on the other. To discuss the nature of the step by which birds took wings and rose out of reptiledom, is to raise the whole problem of organic progress. To refer the step, as Buffon did, simply to a change in the quantity of carbonic acid gas in the atmosphere, is to give the process a false simplicity. To assume the accumulation of reptilian variations by natural selection in the struggle for existence, is the consistent Darwinian view. It is probably more correct to regard the incipient birds as, so to speak, 'fevered representatives of reptiles progressing in the direction of greater and greater constitutional activity.' Flight would be at first spasmodic, but as the structures became modified by function, would gradually become more constant and less pathological. The high temperature, the feathered skin, the flight, the hollow bones, the long rest of brooding, are probably all expressions of one predominant constitutional tendency. It must also be noted that the sacrifice of parental care justifies itself in the success of the brood. See EVOLUTION.
Economic Importance.—In their connection with other parts of the web-like world of life, as well as for their own sakes, birds are of much practical importance. Though eagles are sometimes destructive of lambs, the birds of prey more than make up for this by keeping down the vermin; though many birds rob fruit-trees, devour seeds, and pillage growing crops, their services in keeping away plagues of insects more than counterbalance the damage they do. That some (e.g. humming-birds) visit flowers and act probably as pollen-carriers, and that others aid in the dispersion of seeds, are minor offices. That the flesh and eggs of many, both wild and more or less domesticated, are common articles of food; that the down and feathers of some are in common use for comfort and decoration; that the excrement of others forms the useful manure known as guano, are facts familiar to all. In exceptional cases, birds may be of more curious utility, a variety of pigeon discharges the functions of a letter-carrier, falcons used to be much employed in sport, the nests of one form are eaten, and some birds are kept for amusement and decoration. And when we think of their beauty and song, we must allow that human life would be poorer without them.
Conclusion.—In many centuries, and now more than ever, birds have been studied by the naturalist. The perfection of their structure, the variety and ingenuity of their habits, and in later days the fact of their lofty ascent from lowly parentage, have afforded material for thousands of researches which, in a pre-eminent degree, have been labours of love. In the eyes of simpler men, unacquainted with the technicalities of science, the majority of birds have for long seemed the most delightsome of animals. Apart from their importance as beneficial or hurtful, apart from matters of food and sport, and apart from their ancient augural significance, birds have been universally allowed to possess peculiar charms. Their ethereal habit, their general vivacity, their grace of structure, the beauty of their plumage, the music of their song, the range of their intelligence and emotions, the mystery of their migrations, and other features, combine to make birds the first and last favourites of man. To the artist too they have supplied a wealth of colour, a variety and gracefulness of form, a suggestive beauty of pose and attitude which are not elsewhere equalled. Nor is it necessary to say that just as some of the greatest ornithologists have been at the same time poets, so most poets have been also ornithologists, or at least bird-lovers. To poets of every age and country birds have been 'symbols and suggestions;' although to too many they have been nothing more. They are emphatically the poet's animals; still he is often ignorant of their habits, according to Phil Robinson's indictment in The Poets' Birds (1883).
Literature.—Of the immense literature on birds, the following may be noted:
E. Alix, Appareil Locomoteur des Oiseaux (for mechanism of flight); Balfour's Embryology, vol. ii. (for development); Brehm's Bird Life, &c. and Book of Birds; Brewer, North American Oology (for eggs); British Museum, Catalogue of Birds; Bronn's Thierreich; Cassell's Book of Birds and Natural History (Martin Duncan); Darwin, Descent of Man, &c. (for sexual selection); Foster and Balfour, Elements of Embryology (for development); Gould's Birds of Australia (for reference); Gray and Mitchell, Genera of Birds (for reference); Hewitson, Eggs of British Birds; Huxley, Anatomy of Vertebrate Animals (for structure and classification); Ibis: Journal of Ornithology; Jerdon's Birds of India (for reference); Leunis, Synopsis des Thierreichs (for genera); Macgillivray's British Birds (for anatomy and genera); A. Newton, 'Birds,' Ency. Brit. (for distribution, &c.); Owen, Anatomy of Vertebrates (for structure); W. K. Parker, 'Birds,' Ency. Brit. (for structure and development); J. G. Romanes, Animal Intelligence, Mental Evolution of Animals; H. Seebohm, A History of British Birds, with coloured illustrations of their Eggs, vol. i.-vi. (1886); A. R. Wallace, Contributions to Theory of Natural Selection, Philippine Islands, &c. (geographical distribution of animals); J. G. Wood, Homes without Hands, &c. (for nests and general life); Yarrell's British Birds.