Seaweeds, a general and popular term applied to a vast collection of lower plant-forms growing on the seacoast from high-water mark (or a little above that limit) to a depth of from 50 to 100 fathoms (rarely deeper), and all belonging to the sub-class of the Thallophyta, to which the name Algae has been given (see ALGÆ). Any detailed treatment of the organisms included under the term would be impossible within the limits of a short article like the present. It must suffice, on the one hand, to indicate the chief variations in structure and life-history of typical representatives, and on the other to refer briefly to the more important points in the distribution and classification of the group. The short bibliography appended will supply a guide to the very extensive literature of the subject.
An examination of an average shore-belt exposed at ebb-tide reveals the fact that this area may be roughly divided into four regions: (a) a zone at and above high-water mark, characterised by the predominance of filamentous and encrusting, often inconspicuous, forms, exhibiting a bluish-green colour when examined under the microscope; (b) a zone from near high-water mark to half-tide level, occupied chiefly by algae of a bright grass-green tint; (c) from half-tide level to low-tide mark, where the majority of the plants are olive-brown in colour; and finally (d) from near low-water mark to depths never exposed by the lowest ebb, where the preponderance of forms are of a distinctly red colour. Although not a few shores exhibit these zones fairly clearly, it must be remembered that, owing to the nature of the shore, the character of the tides, and the absence or presence of large influxes of fresh water, the zoning is not always clearly defined. For instance, the uppermost zone of blue-green forms is most likely to be met with on low-lying coast-lines where the seashore proper merges gradually into brackish marshes and muddy pools, such as, for example, the estuaries of the Dee and Mersey, or of the Clyde near Dumbarton. Grass-green forms are most plentiful on shelving rocky shores; whilst on such steep rocky shores as those of the fjords of the west coast of Scotland olive-brown seaweeds form the prevailing vegetation right up to high-water mark. It must be also borne in mind that certain species of algae are invariably found in regions of the littoral area not characteristically occupied by the majority of forms of a similar hue. Thus the common dwarf species, Pelvetia canaliculata, closely allied to the familiar bladder-wrack (Fucus vesiculosus), belongs to the olive division of seaweeds, and yet is invariably found at or even above high-water mark. Tangle (Laminaria digitata), also an olive seaweed, is, on the other hand, a deep-water form only exposed at low tides, whilst the genus Struvea, one of the pure-green algae, may be dredged from 30 fathoms. Very many red algae are found under cover of the olive forms between tide marks, whilst some have their habitat at or above high-water mark. For example, two species of the genus Rhodochorton form a crimson velvety pile on rocks from half-tide level to far above high-water mark, and another and easily recognisable form, Catenella opuntia, grows on the lee side of rocks which are seldom touched by the flood-tide.
It is worthy of note that the classification of seaweeds into four groups according to colour is strikingly supported by the morphology and life-history of the forms so brought together. This fact becomes all the more remarkable when it is remembered that colour among higher plants is in very few cases of even specific value in classifica- tion. The blue-green algae are known as Cyanophyceæ, the pure-green as Chlorophyceæ, the olive as Phæophyceæ, and the red as Rhodophyceæ. (By some authors the lower members of the first two groups are classed together under the name of Protophyceæ.) All possess the green colouring matter chlorophyll, but in the blue, olive, and red forms additional colouring matters (phycocyanin, phycophæin, and phycoerythrin) are present to a greater or less extent, masking the pure-green tint so well seen in the Chlorophyceæ. It is impossible in the present state of our knowledge to dogmatise on the precise value of these additional pigments, but we cannot be far wrong in saying that they are associated with the modification of the intensity or quality of sunlight, and aid or protect the chlorophyll in the peculiar and vitally important duties which it performs in the nutrition of the organism (see CHLOROPHYLL). It has been ascertained that certain rays of the solar spectrum are more efficient than others in the work of assimilation, and it is worthy of note that these rays are precisely those which are first intercepted in the passage of sunlight into sea-water. Our ignorance of the important physiological problems involved may be estimated when we place against this explanation the fact that Kjellman in his exploration of the flora of the Arctic Sea found that algae grew and reproduced at a mean temperature of . and during the long and dark arctic night of three months' duration.
Turning from the bathymetric distribution of seaweeds to their surface distribution, we find here also many interesting and difficult problems. In the first place the medium in which seaweeds live is of a more uniform temperature than that to which land plants are exposed, although against this we must place the fact that seaweeds are more susceptible to fluctuations of temperature. Whilst ocean currents are undoubtedly the chief agents in the transport of the marine flora, long tracts of deep ocean must prove serious barriers to the migration of littoral species. It is scarcely necessary to point out that continental areas, hotter and colder regions of the sea, and long stretches of sandy shore must also act as barriers to possible migration. The effects of such barriers are well seen in comparing the floras of the tropical Atlantic and of the Indian Ocean, the north and south temperate Atlantic, and the eastern and western shores of the same great ocean. From the following table (abstracted from Murray's paper on the 'Distribution of Marine Algae,' Trans. Biol. Soc. Liverpool, vol. v. p. 164) it will be seen that,
| Arctic Sea. | West Indies. | Australia. | Algae common to | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Arctic Sea and West Indies. | West Indies and Australia. | Arctic Sea and Australia. | All three regions. | |||||||||||
| Gen. | Spec. | Gen. | Spec. | Gen. | Spec. | Gen. | Spec. | Gen. | Spec. | Gen. | Spec. | Gen. | Spec. | |
| Rhodophyceæ .. | 44 | 104 | 85 | 444 | 151 | 743 | 21 | 18 | 70 | 79 | 22 | 6 | 17 | 5 |
| Phæophyceæ .. | 42 | 92 | 25 | 112 | 52 | 209 | 9 | 4 | 19 | 21 | 11 | 8 | 6 | 1 |
| Chlorophyceæ .. | 19 | 54 | 30 | 187 | 34 | 149 | 8 | 8 | 15 | 32 | 5 | 7 | 5 | 6 |
| Protophyceæ.... | 6 | 9 | 10 | 45 | 18 | 31 | 4 | 0 | 5 | 3 | 4 | 0 | 4 | 0 |
| Aggregate .. | 111 | 259 | 150 | 788 | 255 | 1132 | 42 | 30 | 109 | 135 | 42 | 21 | 32 | 12 |
taking three principal regions into consideration, the Arctic marine flora consists of 259 species belonging to 111 genera, the West Indian region possesses 788 species and 150 genera, the Australian 1132 species in 255 genera, and that nevertheless only 17 genera and 5 species of Rhodophyceæ, 6 genera and 1 species of Phæophyceæ, 5 genera and 6 species of Chlorophyceæ, and 4 genera and no species of Protophyceæ are common to all.
The structure, life-history, and classification of the lower algae (both fresh-water and marine) having been dealt with in the article Algae (q.v.), there is left for treatment the two higher groups—viz. Phæophyceæ and Rhodophyceæ. It will be most in accordance with the aim of the present article to sketch very briefly the main lines of classification, and to select a few typical examples for more detailed notice.
The Phæophyceæ include all the olive-brown seaweeds found on our shores, and are subdivided according to their methods of reproduction into three chief series—viz. (a) the Phæosporæe, represented by such forms as Ectocarpus, Cutleria, Sphacelaria, and the giants among seaweeds,

A portion of the thallus with multicellular sporangia ().

A, Laminaria digitata, var. stenophylla; B, C, L. digitata, var. cloustoni; D, young form of B (all much reduced). (After Luerssen.)
Laminaria, Macrocystis, and Lessonia; (b) the Fucaceæ—e.g. Fucus, Ascophyllum, Pelvetia, &c.; and (c) the Dictyotaceæ—e.g. Dictyota and Padina. The members of the Phæosporææ have a most varied vegetative form. Many are filamentous and branched, such as Ectocarpus and Sphacelaria, some are tubular and unbranched, as Asperococcus and Scytosiphon, or tubular and branched—Chordaria. Others are ribbon-shaped—e.g. Cutleria; not a few are membranous and encrusting—e.g. Ralfsia; whilst Laminaria forms resemblance to the sieve tubes of Phanerogams, whilst the small cortical cells exhibit that intercommunication of protoplasm which is of so great physiological importance in the higher plants.
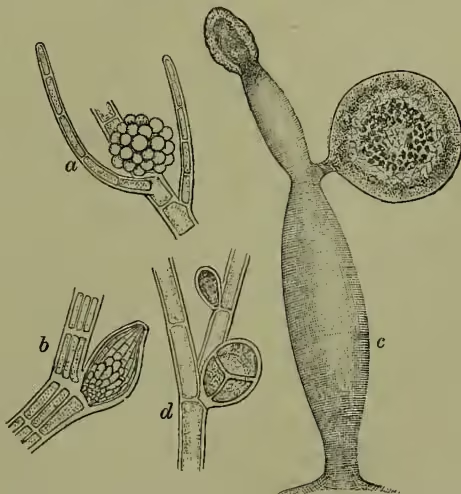
Both sexual and asexual methods of reproduction are known to occur, though not as yet in all genera. Vegetative propagation by gemmæ is also not uncommon. The asexual reproductive organs are in the form of unilocular sporangia, the contents of which are transformed into zoo- spores, small motile cells furnished with two flagella each, and capable after a period of activity of longer or shorter duration of settling down and forming new plants. The sporangia either are developed externally on the thallus or are the terminal cells of short branches. They are frequently collected in groups (sori), and are accompanied by sterile filaments. The sexual organs are multicellular sporangia (gametangia), of diverse form, each locus giving rise to a single motile cell somewhat like a zoospore. These cells conjugate in pairs, the product (zygote) behaving in a precisely similar manner to the zoospore. Although in most of the Phæosporææ the zoogametes are morphologically indistinguishable, interesting gradations may be observed in some genera where the zoogametes are differentiated into male and female cells. Cutleria is especially interesting in this respect, as the female gametes are much larger than the male gametes, and come to rest before conjugation with the still motile male cell.
The Fucaceæ are characterised by having distinctly differentiated sexual organs with non-motile ova and motile fertilising cells. The ova are formed within the swollen cells (oogonia) which arise from the wall of flask-shaped cavities (conceptacles), and are accompanied by unbranched hairs. The fertilising cells, antherozoids or sperms, are produced in ovoid terminal cells of branched hairs (antheridia), which likewise arise from the walls of similar conceptacles. In Fucus platycarpus both ova and sperms are formed in the same conceptacle; in most Fucaceæ, however, they are formed on different plants. Asexual

A portion of the thallus showing receptacles (). (After Thuret.)
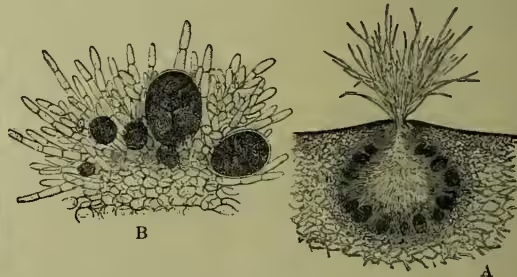
A, vertical section through a conceptacle; B, portion of the wall of a conceptacle, showing oogonia in various stages of development, and antheridia. (A, ; B, .) (After Thuret.)
multiplication is unknown. The group includes such well-known genera as Fucus, Halidrys, and Pelvetia, with branched leathery fronds, the tips of whose branches become transformed into receptacles for the reproductive organs, and Himanthalia, with a button-shaped thallus, from which arise the long strap-shaped reproductive branches characteristic of that genus.
The Dictyotaceæ are an aberrant group of Phæophyceæ with distinct affinities to the Rhodophyceæ. They resemble that group in having non-motile spores produced in fours in sporangia which are clustered in sori over the surface of the frond (Dictyota) or along the midrib. The sexual organs are in the form of antheridia and oogonia. The male organs produce non-motile fertilising cells resembling the pollinoids of the Rhodophyceæ, though Johnson (Jour. Linn. Soc. Bot., vol. xxvii. p. 467) considers that possibly the pollinoids of Dictyopteris may be ciliated like those of Cutleria. The oogonia are arranged in sori, each containing one ovum. In both Fucaceæ and Dictyotaceæ fertilisation is external.


The Rhodophyceæ (Florideæ) form a very large assemblage of most varied vegetative form, and every possible shade of red from a purple black to brilliant pink. The root may be a branched mass, a plate, or a disc attached to mud, other algæ, or rock, whilst the fronds are filamentous, membranous, crustaceous, or calcareous. The asexual organs consist of sporangia whose entire contents in some genera escape as a single non-motile spore. In the majority, however, each sporangium contains four non-motile spores. The sporangia are solitary or grouped in sori, and often sunk in the tissue of the frond or in special branches. The sexual organs are antheridia and procarpia. The antheridia are generally modified terminal branches or special areas in the thallus (of the more succulent forms), from which are derived short rod-like fertilising cells, here called pollinoids. The female organ is a procarp, whose structure varies in complexity in the different orders of Rhodophyceæ. In the simplest condition it recalls the female organ of Coleochaete, and consists of a rounded cell with a long filamentous apical portion, the trichogyne. In higher members of the series the trichogyne is separated from that portion of the procarp from which the products of fertilisation (carpospores) are derived by one or more intermediate cells, the trichophore, and the basal part itself may become multicellular, all or only some of the cells of which produce the pro- ducts of fertilisation. Fertilisation is effected by fusion of a pollinoid with the trichogyne, the immediate result being the formation of a cluster of cells, the carpospores, derived from the fertile cells of the base. These carpospores may form a mulberry-like mass (Nemalion), or be enclosed by a loose involucre of filaments (Griffithsia), or by an ovoid capsular investment with a terminal pore (Polysiphonia). In others again the collection of carpospores (cystocarp) is completely enclosed within a thick-walled spherical sac (Plocanium), or may be sunk in the tissue of the frond (Dumantia). The Rhodophyceæ embrace a large number of orders, the representatives of several of which form well-known objects of interest on the seashore. The best known of these perhaps are Carragcen (Chondrus crispus), Dulse (Rhodymenia palmata), and Laver (Porphyra laciniata). The genera Callithamnion, Delesseria, Nitophyllum, Plocanium, and Odonthalia are well known to collectors for the beauty of their fronds. In concluding this article it may be of interest to point out that the vast collection of feathery plant-like
Fig. 7.—Corallina officinalis, a calcareous seaweed (natural size).
forms, popularly classed as 'seaweeds' by seaside visitors and sold as such after being fancifully tinted, are in no respect related to the forms we have been considering, but really belong to the zoophyte division (Hydrozoa) of the animal kingdom.
For British Seaweeds, see Harvey, Phycologia Britannica; Gray, British Seaweeds; Landsborough, Popular History of British Seaweeds. General and Systematic: Agardh, Species, Genera, et Ordines Algarum; Thuret, Études Phycologiques; Bornet et Thuret, Notes Algologiques; Hauck, Die Meeresalgen; Reinke, Atlas Meeresalgen; Falkenberg in Schenk's Handbuch der Botanik; Schmitz, Untersuchungen über die Befruchtung der Floriden.