Circulation, in Anatomy and Physiology, is the term used to designate the course of the blood from the heart to the most minute blood-vessels (the Capillaries, q.v.), and from these back to the heart. To simplify the consideration of the subject, we shall consider—(I.) the General Anatomy, (II.) the Comparative Anatomy, (III.) the Physiology, and (IV.) the History.
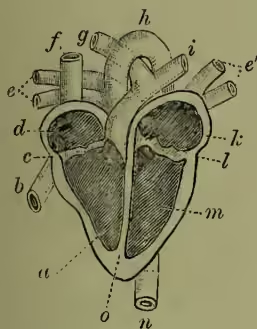
I. The organs of circulation consist of the heart, arteries, veins, and capillaries. The course of the blood through these organs will be best elucidated by the aid of a diagram, which is equally applicable for all other mammals as well as for man, and for birds. The dark parts of fig. 1 represent the course of the impure or venous blood, while the lighter portions represent the course of the pure or arterial blood. Two of the four chambers of the heart (A and C) receive the inflowing blood, and are termed the auricles; while the other two chambers (B and D) drive the blood to the lungs and to the general system respectively, and are termed the ventricles. The vessels that bring blood to the auricles are termed veins, and the vessels through which the blood is driven from the ventricles are known as Arteries (q.v.). We will now trace the course of the blood, as indicated by the arrows of the diagram, commencing with the right auricle, A. The right auricle contracting upon the venous or impure blood with which we suppose it to be filled, drives this into the right ventricle, B, through an opening guarded by a triple (or tricuspid) valve, which almost entirely prevents the regurgitation of the blood from the ventricle into the auricle. The ventricle, B, being now filled, contracts, and as the blood cannot return into the auricle, it is driven along the dark vessel, c, which is the pulmonary artery, conveying the blood to the lungs. At its commencement it is guarded by crescent-shaped (semilunar) valves, which entirely prevent the blood which has once been propelled into the pulmonary artery from re-entering the ventricle. The pulmonary artery gradually divides into smaller and smaller branches, which ultimately emerge into capillaries freely distributed over the interior of the air-cells of the lungs. There the blood gives off carbonic acid gas, and absorbs oxygen, becoming thereby 'pure.' The capillaries, in which the blood is purified, gradually unite to form larger vessels, and finally the blood is collected into the pulmonary veins, which pour their contents into the left auricle, C. Thence it is propelled into the left ventricle, D, through an opening guarded by a double (mitral or bicuspid) valve, which entirely prevents the reflux of the blood. The left ventricle contracts and drives its contents into the large artery, e, or Aorta (q.v.), which by means of its various branches supplies the whole body with pure blood. From the aorta and its various subdividing branches the blood passes into the capillaries, J, H, which occur in every part of the system. In these capillaries it parts with its oxygen to the tissues, and becomes charged with the waste carbonic acid gas. The capillaries unite into larger veins, and these gradually unite to form two large trunks, a, b, the superior and inferior venæ caveæ, which pour their contents into the right auricle—the point from which we started. The diagram also shows how the venous blood from the viscera, G, instead of passing directly into the
Fig. 1.—Diagram of Circulating System (from Leunis).
vena cava, goes through the liver, F, in what is known as the hepatic-portal system.
Before passing to the comparative study of the circulation, we must notice a few of the above facts in greater detail.
The Heart (q.v.) is described in a separate article. Since all the arterial blood leaves the heart through the aorta, to trace the circulation of the pure blood involves following the branches of that great vessel. For this see AORTA. It is sufficient, without further anatomical details, to say that the ramifications of the arteries distribute the arterial blood to the Capillaries (q.v.) which pervade every part of the body. It is already evident that the pulmonary arteries from the right ventricle carry impure blood to the lungs, and that pure blood returns from the lungs to the left auricle by the pulmonary veins.
The veins, like the arteries, are found in nearly every tissue; they commence in minute networks which communicate with the capillaries. Branches from these networks uniting together, form veins, which, by joining, increase in size as they pass onward towards the heart. If we except certain venous structures (called sinuses) occurring in the interior of the skull, we may divide the veins into two sets—the superficial or cutaneous, and the deep veins. The deep veins accompany the arteries, and are usually inclosed in the same sheath of tissue. In the case of the smaller arteries they generally exist in pairs, one on each side of the artery, while the larger arteries have usually only one accompanying vein. The superficial veins occur immediately beneath the integument; they not only return the blood from the skin and adjacent structures, but communicate with the deep veins. All the veins finally lead by two large trunks, the superior and inferior vena cava, into the right auricle of the heart. The superior vena cava is formed by the union of the veins from the head and neck (the jugulars) with those from the arms (the subclavians), while the inferior vena cava brings back the blood from the lower extremities, the trunk, and the viscera.
We must refer to the article VEIN for the structure of the walls of this part of the circulatory system. There is only one point that imperatively requires notice here—viz. that while the arterial system presents no valves except at the points where the two great trunks leave the heart, the veins contain a great number of valves, which are formed by a doubling of their lining membrane, and resemble pocket-like folds or pouches, which allow the blood free passage toward the heart, but prevent its reflux. The veins are much less elastic than the arteries, and their total capacity is much greater.
There is one part of the venous circulation which, from its great importance, requires special notice —viz. that of the spleen, pancreas, stomach, and intestinal canal. The blood supplied to these organs by the cœliac and the two mesenteric arteries is not returned directly to the inferior vena cava, but passes by several veins into one large vessel—the portal vein, which enters the liver, and breaks up into a capillary network. There the blood undergoes important changes associated with the bile-secreting and glycogen-forming functions of the liver. The blood, entering the liver from two sources, from the portal vein and from the hepatic artery, leaves it by the hepatic veins, which join the inferior vena cava. It is also important to notice the entirely distinct set of vessels known as lymphatics, which conduct the products of digestion into the veins (see LYMPH).
The above-described double circulation (through the lungs and through the body) is exhibited by the blood from the time of birth during the whole period of life. The circulation of the blood, however, begins before birth—indeed, at a very early period of intra-uterine or fœtal existence; and the circumstance that before birth the lungs do not act as organs of respiration induces a very important modification in the course of the blood in fœtal life which will be described under FÆTUS.
II. Comparative.—The circulatory system in man, as above described, may serve as type of the highest development, differing but slightly from that of other mammals, or that of birds. It is convenient now to begin at the other end, and to note briefly the salient steps of progress in the gradual evolution of the system throughout the animal series. In the unicellular animals the movement of the protoplasm and the special activity of 'contractile vacuoles,' represent, to some extent at least, a circulatory function before the appearance of any system. The canals which so completely irrigate a sponge, likewise illustrate in low expression a circulatory system not yet separated off from the others. In Cœlenterates, too, the system is still unseparated: 'gastro-vascular' prolongations of the alimentary cavity penetrate the body, as may be very well seen in the disc of a common jelly-fish. In the lower worm-types also, where no distinct body-cavity is yet developed, the nutritive fluid simply diffuses through the body, and no vascular system is differentiated. But in higher worms there is generally a body-cavity, and with it the gradual appearance of a definite vascular system. In some we simply find a fluid moving in the body-cavity, occasionally clear, usually with corpuscles; in others, portions of the body-cavity are separated off as blood-spaces, or eventually as blood-vessels; these may remain in connection with the general cavity, or may at length form a closed system. The manifold worm-types afford abundant illustration of all the stages in this differentiation. In the bristle-footed worms (Chætopods, q.v.), and in some others, the perfecting of the blood-driving mechanism may be instructively traced. Often a dorsal vessel is diffusely contractile, less frequently the ventral; or there may be contractile connecting loops between dorsal and ventral vessels, as in the earthworm; or lastly, a special region in the dorsal vessel may become the main seat of the vascular contractility. Such a dorsal heart is found from this point onwards throughout the Arthropods and Molluscs. (The well-developed and very difficult vascular system of Echinoderms, which co-exists with an abundant body-cavity fluid with relatively few corpuscles, has no special interest for this general survey.) In crustaceans, the dorsal heart, usually inclosed in a special space or 'pericardial sinus,' drives blood by more or less well-developed arteries through the body. The rest of the system is best described as lacunar.
The venous blood passes along body-cavity spaces to the gills for purification, thence returns to the pericardial sinus, and entering the heart is redistributed. In insects a chambered dorsal heart, inclosed in a sinus as before, drives the blood forward, but as one would expect from the very efficient respiratory apparatus, the general vascular system is but slightly differentiated. The blood, purified by diffusion from the everywhere present air-tubes, passes back by venous channels into the sides of the pericardium and heart. In myriapods, scorpions, and king-crab, the system is more definitely developed, but illustrates no new advance except that of more complete establishment and wider extension of vessels. In molluscs, however, some progress is observable. Except in the Elephants' Tooth Shell (Dentalium), a heart is present, and the arterial system is often very well developed, even to the extent of capillaries in some cuttle-fishes and snails. Usually, however, the venous blood travels along lacunæ, though gradual transitions occur between these and true veins. The blood purified in the gills or pulmonary chamber passes back into a special portion of the body-cavity—the pericardium, and thence into the heart. In certain worm-types, several contractile lateral vessels may often be observed to enter the dorsal vessel; in the Pearly Nautilus, which has four gills, four efferent vessels dilating into four indistinct auricles, enter the median dorsal heart or ventricle; in almost all bivalves the entrant dilatations or auricles are reduced to two, one on each side; while in most Gasteropods and Pteropods the specialisation has gone further, and the heart consists of a single auricle and a thicker muscular ventricle. The latter drives the blood through the body by a single or double aorta.
Passing now with equal brevity through the vertebrate series, we notice first that the heart arises as a dilatation no longer of a dorsal, but of a ventral vessel. Up to and including amphibians, the heart begins as a specialisation of the 'sub-intestinal vein' in the throat region; in most, if not all higher vertebrates, it arises from the fusion of two vessels. It always lies in a pericardial sac.
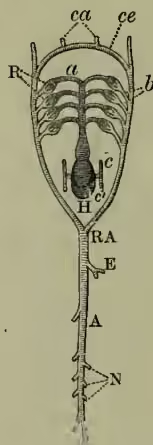
(1) Among the degenerate Tunicata there is considerable variety in the vascular system. In one case no heart is present; in several there are no definite vessels or blood-corpuscles. The main point, however, is that in most cases a tubular ventral heart drives blood to the respiratory pharynx. In all cases where the heart has been observed, the direction of its beats has been seen to undergo reversal at regular short intervals, a phenomenon which has also been noticed as a rarity in certain worm-types.
(2) The vascular system of the lancelet or amphioxus is of a peculiarly diffuse and undifferentiated nature. It has in one sense no heart, in another sense many; for while there is no main centre of contractility, there are small pulsating dilatations at the bases of the vessels passing to the gill-slits, while the portal vein and ventral vessel in the anterior pharyngeal region are both said to be contractile. In general course, the circulation is like that of a fish; the blood passes from ventral vessel to respiratory region, thence to dorsal aorta, thence to body, thence by united sub-intestinal veins to the liver cæcum, and thence to the ventral vessel from which it started.
(3) In the Round Mouths (Cyclostomata) the typical fish-circulation is established. The muscular ventricle drives the blood by a ventral vessel ('ventral aorta') to the gill-sacs; thence the purified blood is gathered into efferent dorsal vessels, which form in uniting the 'dorsal aorta.' The latter gives off branches to the greater part of the body, the head-region being directly supplied from the anterior efferent branchials. The blood returns from the anterior and posterior regions into a uniting vessel behind the heart ('the sinus venosus'), thence into the receiving auricle, and from that to the muscular ventricle.
(4) It is enough after the above to notice in regard to the fishes proper, that with the exception of the double-breathing mud-fish (Dipnoi), the heart never contains anything but impure blood, that it drives this wholly to the respiratory organs, and is in no direct degree 'systemic.' The 'dorsal aorta' supplying most of the body is formed from the union of efferent branchials, and does not arise, as in higher vertebrates, from the heart. It is important to notice that the five or so arches which spring from the ventral aorta are almost all quite alike, and arise (except in Dipnoi) at slight intervals from one another. A great part of the differentiation in higher vertebrates obviously concerns these aortic arches, which are seen in Cyclostomata and fishes in primitive uniformity, but become modified in higher vertebrates into the carotids, sub-clavians, aortic arches, and pulmonary arteries. The heart of a fish consists of the sinus venosus or general junction, running transversely behind the heart (persisting hence onwards, except in adult birds and mammals), of the auricle and ventricle, and except in Teleostei of a specialised contractile portion of the latter
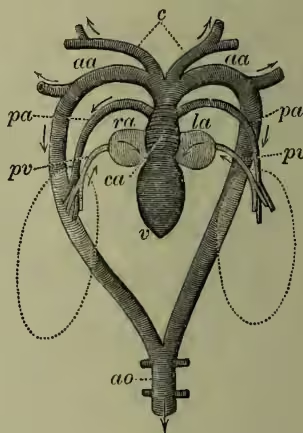
ra, right auricle; la, left auricle; v, ventricle; ca, conus arteriosus; c, carotid arteries; aa, aortic arches; ao, dorsal aorta; pa, pulmonary artery; pv, pulmonary vein; ra receives venous blood from body; both the pulmonary arteries enter la. (After Nuhn.) known as the eonus arteriosus. A dilatation of the beginning of the ventral aorta is distinguished as the bulbus arteriosus. In regard to the general system, it is worth noticing that in fishes (as in amphibians and all reptiles except Chelonians) there is a renal portal, as well as an hepatic portal system. Veins from the caudal region of the fish come into the same relations with the kidneys as the portal veins do in regard to the liver. The hepatic veins returning from the liver do not unite with the other posterior veins, but enter the sinus venosus independently. Thus fishes have no inferior vena cava. (4a) The Dipnoi are interesting as leading on to amphibians. The heart receives pure as well as impure blood, drives blood to lungs in addition to gills, is in part systemic (driving pure blood in Protopterus and Lepidosiren through the first two pairs of arches), has the roots of the aortic arches close together, and is practically three-chambered. In these respects it reaches forward to the condition seen in those amphibians which retain their gills.
(5) The heart of amphibians is three-chambered, a right auricle receiving impure blood from the body, a left auricle receiving purified blood from the lungs or from the lungs and gills, a single ventricle which drives the blood, mingling less than might be supposed, to head or body or respiratory organs. In some cases, as one would expect, the partition between right and left auricle is imperfect. In the tadpoles at the fish-like stage there are four aortic arches, of which the first three supply gills. Where gills persist, such supply is always of course in some degree maintained. In such a metamorphosis as that of the frog, where all trace of gills is lost, the first branchial arch becomes the carotid; the second is the systemic, which forms, by uniting with its fellow, the dorsal aorta; the third dwindles away; the fourth supplies the lungs. The same is generally true of the higher vertebrates, except that it is usually the third branchial which forms the pulmonary artery. An inferior vena cava is definitely established in amphibians; there is a renal portal as well as an hepatic portal system; special lymph hearts are sometimes present connecting the lymph system with the vascular.
(6) Among the reptiles, the differentiation of the heart goes a step further. In Chelonians, Lacertilia, and Ophidia, a strong muscular ridge forms an incomplete partition, dividing the ventricle into a right portion containing purely venous blood, in the ventricle, and thus for the first time a four-chambered heart, though it is not certain that the division is exactly comparable to that of birds and mammals. From the right or venous ventricle rise the pulmonary artery and left aortic arch (taking venous, not mixed, blood to the viscera); from the left or arterial ventricle rises the right aorta, with pure blood to the greater part of the body. Though the division of arterial and venous chambers brings the crocodilian heart to the bird or mammal level, there are two aortic arches, one venous, the other arterial, which unite as usual, and also communicate by a foramen at their roots. In all reptiles there are two superior venæ cavae, an inferior vena cava formed from the union of the two efferent renals, and a renal portal system except in Chelonia. In birds and mammals a single aortic arch forms the dorsal aorta, not two or more as in reptiles. In birds the aorta goes to the right, in mammals to the left, but except in this variation and in the structure of the Heart (q.v.), the circulation in Birds (q.v.) and mammals (supra) is practically identical.
III. Physiology.—The most important conditions of circulatory function are (1) a general constancy and rapidity of flow, and (2) a power of adapting this to special needs. The conditions of the former are mainly mechanical and physical—those of the latter depend upon the nervous system.
(1) The Mechanism of Circulation chiefly depends upon the rhythmic contractility of the heart, the elasticity of the vessels, and the friction in the small arteries and capillaries. The heart is a muscular pump contracting under the influence of its automatic nervous mechanism, but also responding in the nature of its beat to the conditions of the body. The human heart usually beats about seventy-two times a minute, and in the eight-tenths of a second occupied by each beat, three distinct events occur—the contraction (systole) of the ventricles, a 'passive interval' of relaxation (diastole) of both auricles and ventricles, and the decidedly briefer contraction of the auricles. The pumping action of the heart is thus obviously an intermittent force which drives the blood through the closed series of elastic tubes formed by the blood-vessels. To the latter attention must now be directed.
The arteries are surrounded by muscular tissue, and are contractile as well as highly elastic. As they break up into branches from the aorta to the capillaries, the capacity for holding blood is continually increasing. The flow of blood within them, though continuous, comes in gushes corresponding to the heart-beats. The velocity of the stream is greatest the nearer the heart. The veins are much less elastic than the arteries, and have a much greater total capacity for holding blood. As they unite from the capillaries to the venæ cavae, the total capacity is continually diminishing. The flow of blood within them is continuous, but with relatively little force and small velocity. The capillaries have a very small calibre, but are both elastic and contractile. They are permeable by fluids, and also allow the corpuscles to pass through their walls. The velocity of the flow is here at its minimum. The resistance to the flow of blood, due to the friction of these minute passages, works back along the arterial system to the heart, and is one of the most important factors in determining the nature of the circulation.
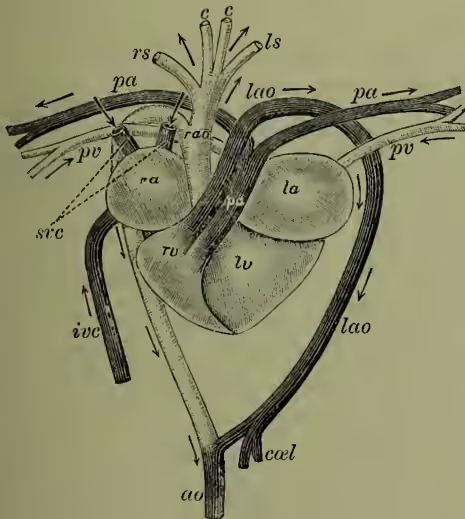
To return now to the actual mechanism, it is only necessary to note (a) that the whole system is always over-filled with blood, which consequently causes a pressure on the walls; (b) that the cause of the circulation is the difference of pressure between the blood in the aorta and pulmonary arteries on the one hand and the venæ cavae and pulmonary veins on the other; (c) that this difference of ra, right auricle; la, left auricle; rv, right (venous) portion of ventricle; lv, left (arterial) portion of ventricle; lao, left (venous) aortic arch; pa, pulmonary artery; rao, right (arterial) aortic arch; rs, ls, branches to fore-limbs; cc, carotids; pv, pulmonary veins; sve, superior venæ cavae; ivc, inferior vena cava; ao, dorsal aorta; ca, celiac artery to viscera. (After Nuhn.) and a left portion containing mixed and arterial blood. The pulmonary artery rising from the right cavity takes purely venous blood to the lungs; of the two aortic arches rising from the left cavity, the left aorta contains more venous than arterial blood, the right aorta more arterial than venous. In Crocodilia, however, there is a complete septum pressure is due to the contraction of the heart; and (d) that the interrupted current, which would naturally arise from the intermittent action of the heart, becomes continuous because of the rapidity of the beats, the resistance in the small arteries and capillaries, and the elasticity of the arterial walls.
(2) The Control of the Circulation.—The phenomena referred to above are mainly physical and mechanical, and in their essential features can be readily reproduced on a model. But in a complex living organism there is a constant necessity for modifiability. The organs vary in their requirements, and the external conditions are frequently changing. Modifications in the circulation are brought about by changes in the heart's beat, and in the calibre and resistance of the small arteries. Both these changes are under the direct control of the Nervous System (q.v.). Modifications of less importance also occur in the capillary resistance and in the total quantity of blood; but the important point is simply the general fact that the requirements of the organism are met by the dominion of the nervous system over the circulation.
IV. History.—Aristotle and the ancients thought of the heart as a blood-fountain, but had no conception of circulation. Galen (131-201) disproved the prevalent notion that the arteries contained air, Calvin's contemporary Servetus demonstrated the pulmonary circulation, and various physiologists had a distinct notion of the centrifugal flow. In 1628 William Harvey published his Exercitatio Anatomica de Motu Cordis et Sanguinis in Animalibus, in which his discovery of the real nature of the circulation was expounded. Soon after, the use of the microscope revealed to Malpighi (1661) and Leuwenhoek (1674) what Harvey had been unable to trace—the capillary connection between arteries and veins. Since those days the detailed problems of circulation, the hydraulic mechanism, the action of the heart and its valves, and the nervous control have been worked at by many of the greatest physiologists.
See AORTA, BLOOD, HEART, VEIN, &c.; the physiological text-books of Foster, Hermann, Huxley, Landois, Stirling, &c., for human circulation; works on comparative anatomy (Jeffrey Bell, Claus, Gegenbaur, Huxley, Wiedersheim, &c.), for the gradual evolution of the system; Willis's William Harvey, for the earlier notions on the subject.